
Rhinopithecus bieti Milne-Edwards, 1897
|
publication ID |
https://doi.org/ 10.1093/mspecies/sey016 |
|
publication LSID |
lsid:zoobank.org:pub:4D9C0C1A-B712-4246-84F9-18CAE8EC6272 |
|
DOI |
https://doi.org/10.5281/zenodo.4605126 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE2B5D-FFD4-8D4F-FF54-9D84FBF6FCA5 |
|
treatment provided by |
Felipe |
|
scientific name |
Rhinopithecus bieti Milne-Edwards, 1897 |
| status |
|
Rhinopithecus bieti Milne-Edwards, 1897 View in CoL
Black-and-white Snub-nosed Monkey
Rhinopithecus bieti: Milne-Edwards, 1897:156 View in CoL . Type locality: “...aux environs de Tsékou et à Atentsé dans la haute vallée du Mékong” (around Tsékou and Atentsé [modern Yunling Xian, 98.86° E, 28.29° N] in the upper valley of the Mekong), Yunnan, China. First use of current name.
Rhinopithecus roxellanae bieti: Ellerman and Morrison-Scott, 1951:202 View in CoL . Name combination.
Pygathrix (Rhinopithecus) roxellana bieti: Groves, 1970:569 View in CoL . Name combination.
Rhinopithecus (Rhinopithecus) bieti: Jablonski and Peng, 1993:36 View in CoL . Name combination.
CONTEXT AND CONTENT. Context as for genus. Rhinopithecus bieti is monotypic.
NOMENCLATURAL NOTES. Milne-Edwards (1870:341) 1st described Rhinopithecus roxellana , placing it in the genus Semnopithecus of Dufresne (1797). He later changed the genus to Rhinopithecus and the species name to roxellanae (Milne-Edwards 1872:158) . Anderson (1878:43) listed Semnopithecus (Nasalis) roxellana among Yunnan taxa but on his expeditions did not see or obtain specimens of any Rhinopithecus species.
Milne-Edwards (1897:121; pls. IX–XII) did not designate a type specimen in his original description of R. bieti , but described, illustrated, and gave measurements for a “mâle vieux” among his 7 specimens for which Elliot (1913:104) repeated measurements, referring to this mature male as the “Ex type ” in the Paris Museum. Allen (1938:302) concurred that this specimen “might be selected as the type.”
Pocock (1934) had “created a genus Presbytiscus for it [ R. avunculus ], a taxon which seems altogether unnecessary...” ( Groves 1970:559). Ellerman and Morrison-Scott (1951) had earlier considered Presbytiscus valid as a subgenus, although it was often used as the genus untilJablonski and Peng (1993) confirmed it as a subgenus: R. (Presbytiscus) avunculus .
Chasen (1940) followed Elliot (1913) and Allen (1938) in maintaining R. roxellana , R. bieti , and R. brelichi as species of Rhinopithecus , but Ellerman and Morrison-Scott (1951) reduced R. bieti and R. brelichi to subspecies of R. roxellana . Later, Ellerman and Morrison-Scott (1955:19), noting contra Chasen (1940) that “…it appears unnecessary to have three genera for three snub-nosed Monkeys,” placed Simias , Presbytiscus , and Rhinopithecus as subgenera under Rhinopithecus , but did not name the species and subspecies as such.
Groves (1970), Szalay and Delson (1979), and others placed the 4 species of Rhinopithecus known at that time in a subgenus Rhinopithecus under Pygathrix , but as information on their distinctiveness accumulated, Ye et al. (1985) followed by Peng et al. (1988) and Groves (1989) accepted R. bieti as a full species. Nevertheless, Delson (2000) and others continued to recognize Pygathrix (Rhinopithecus) . Although Jablonski and Peng (1993) resurrected Ellerman’s and Morrison-Scott’s (1951) subgenus: R. (Roxellanae) bieti, Groves (2001) renamed it R. bieti .
Geissmann et al. (2010:1) described the 5th species, R. stykeri in Myanmar, near the border with the mainland of China, designating it the Burmese snub-nosed monkey. Its range has since been extended to mainland China west of the Salween River (Nu Jiang) with an estimated population in mainland China of 490–620 ( Ma et al. 2014).
Rhinopithecus bieti is named for Monsignor Biet, apostolic bishop of Tibet, who had arranged for the collection of the original specimens (Milne-Edwards 1897). It has been called the Yunnan or black snub-nosed monkey, but since it also occurs in Tibet (Xizang province of mainland China), Yunnan snubnosed monkey seems restrictive. Since R. strykeri , which has an all-black body, was described in 2010, Chinese scholars have been calling it the black snub-nosed monkey (e.g., Chen et al. 2015), and R. bieti the black-and-white snub-nosed monkey (e.g., Clauzel et al. 2015). Black-and-white snub-nosed monkey in Chinese is “Hei Bai Yang Bi Hou,” Ḇ H ® ⁂ M. Bai people call it “Baiyao” or “Baip hhod”; the latter means “monkey wearing white trousers.” Lisu people call it “Zha mi pu zha” (“Zha mi” means “monkey” in the Lisu language). In Tibetan, R. bieti is “Zhi Jie.”
DIAGNOSIS
Rhinopithecus bieti is 1 of the largest colobines: males’ head–body length is 740–830 cm, tail length is 510–720 cm. Males are 50% larger: 15.3 kg, n = 5 for males, versus 9.1 kg, n = 7 for females (Kirkpatrick 1998). Its tail is 86–95% of head–body length ( Groves 1970). In common with other snubnosed monkeys, it has thick lips, a flat face, and a diminutive nose with upturned nostrils. R. bieti is the only long-haired, black-and-white snub-nosed monkey. Males have a dark crown with hairs 97–120 mm long forming a forward-drooping crest ( Groves 1970). Its facial skin is bare and white with a black nose and pink lips. Pelage is black on the arms (but with a tuft of white at the elbows), back (but with pale guard hairs), and lower legs, whitish-gray on the cheeks, neck, thighs, and underparts, grading to a white abdomen with a dark midline ( Fig. 1 View Fig ). “Hairs very elongated in ischial region forming a fringe 200– 210 mm long” ( Groves 1970:568). White hair on the thighs of mature males is very long and beard-like. Females and immatures are not as dark and their hair is shorter on the thighs, crown, and back. Infants are white and turn yellowish before turning gray. The color pattern is broadly similar to R. avunculus , endemic to Vietnam, which has a white crown, white cheek, and neck hair and is smaller and much shorter-haired, a likely adaptation to the milder climate. R. strykeri has a similar face, but black pelage, and occurs between the Salween (Nu) and Irrawaddy rivers on both sides of the Myanmar–China border ( Ma et al. 2014).
GENERAL CHARACTERS
Rhinopithecus shares many cranial, dental, and postcranial features such as body size and limb proportions with the proboscis monkey, Nasalis larvatus , duoc langurs, Pygathrix , simakobu (pig-tailed langur), Simias concolor , and the fossil Eurasian colobine Mesopithecus , suggesting a common inheritance (Peng et al. 1993; Peng and Pan 1994; Jablonski 1998). Pan et al. (2004) recommended placing these 4 genera and Rhinopithecus into a tribe, the Rhinopithecini . Alternatively, they are referred to as the “odd-nosed” clade, the Rhinopithecus clade, or the Rhinopithecus complex ( Jablonski 1998; Sterner et al. 2006; Whittaker et al. 2006; Roos et al. 2011). These common features contrast with the analogous features of the surilis, Presbytis , lutungs, Trachypithecus , and south-Asian langurs (henceforth, langurs), Semnopithecus , collectively called the Presbytini ( Groves 2001) .
Rhinopithecus bieti is the most sexually dimorphic of snubnosed monkeys for size (as well as pelage differences as noted above) with females 63% to 69% male body mass (Jablonski and Pan 1991, 1995). As well, sexual dimorphism in canine size ( Fig. 2 View Fig ) of Rhinopithecus species is among the highest for colo-bine monkeys, and among species of Rhinopithecus , is highest in R. bieti ; this may be associated with intense male–male compe-tition in the restricted breeding season (Jablonski and Pan 1995). Grueter and van Schaik (2009) showed that colobines living in multilevel societies (with more frequent and intense reproductive competition), as R. bieti does, exhibit higher levels of sexual dimorphism in body mass.
Brachial index is 97–107 and intermembral index is 92–96 for all species of Pygathrix as then understood to include Rhinopithecus ( Groves 1970) . The shoulder gir-dle and upper arm are also more robust in males (Pan et al. 1993), probably reflecting their greater terrestriality ( Kirkpatrick et al. 1998).
Rhinopithecus bieti is primarily arboreal, but with a strong terrestrial component of locomotion and occasional use of alpine habitats above the tree line (Kirkpatrick and Long 1994). Adult males, being heavier, are consistently more terrestrial than females ( Kirkpatrick et al. 1998; Grueter et al. 2013b).
DISTRIBUTION
Rhinopithecus bieti is confined to the Hengduan Mountains between the Mekong River (Lancang Jiang) and the Yangtze River (also named Chang Jiang, “long river”; and Jinsha Jiang in the upper reaches) inYunnan, and Xizang (Tibet Autonomous Region, hereafter “ Tibet ”), mainland China ( Fig. 3 View Fig ). The current range extends from the south near Yunlong (25°52′N) in Lanping and Yunlong Counties, to the north at Zhuina, 29°20′N, Mankang County ( Long et al. 1994). Allen (1938:303) noted that “Prince Henri d’Orleans and M. Bonvalot... in 1890 saw them in the forest country between Tengri Nor and Batang.” This extends the historic range northward close to 30°00″N ( Long et al. 1994). To the south, historical records exist to about 25°12′N (He and Zhang 1992; Li et al. 2002; Nüchel et al. 2018). The southernmost population was thought to be at Mt. Longma at 26°14′N ( Long et al. 1994) in the Yunlong Provincial Nature Reserve until 2003–2007 when staff of Tianchi Provincial Nature Reserve obtained photographs of R. bieti , conducted surveys finding several groups, and collected droppings for food analysis. In February 2015, nature reserve staff captured still and video images of 15–30 individuals at the Wubaoshan (Wubao Mountain) section of the reserve ( Fig. 3 View Fig ), moving the known distribution south about 40 km (L.-X. Han, in litt., 28 April 2015, citing local news reports). Wubaoshan is at 25°52′N, 99°15′E in Yunlong County. These 2 nature reserves are adjacent and are often called the YunlongTianchi Nature Reserve.
A larch-fir forest in the Cang Mountains (Cangshan, west of Dali in Fig. 3 View Fig ), adjacent to the Erhai Basin, contains suitable habitat but is currently unoccupied (He and Zhang 1992; Long et al. 1994). Diancangshan (Dian Cang Mountain, 4,090 m) is at 25°40′N, 100°5′E. Interviews by Dali University in 2014 showed that R. bieti occurred in Cangshan region before 1980s (Zhao et al. 2017).
Contrary to some reports, Long et al. (1994) provided convincing evidence that, despite the appearance of suitable habitat east of the Yangtze River, R. bieti has never occurred there and local people have no knowledge of it. Evidently, the Yangtze and Mekong Rivers have been barriers to dispersal.
FOSSIL RECORD
Mesopithecus pentelucus is a probable ancestor of the snubnosed monkeys (Peng et al. 1993; e.g., Delson 1994; Jablonski 1998; Groves 2000; Ravosa and Profant 2000; Pan et al. 2004; Jablonski et al. 2011). Recently, a Mesopithecus fossil has been found in Zaotong, northeast Yunnan ( Jablonski et al. 2011). Colobines arrived in mainland China in the late Tertiary, later than the Cercopithecines (Pan and Jablonski 1987). Rhinopithecus fossils of Pleistocene age have been found throughout western China and Vietnam (Pan and Jablonski 1987; Jablonski 1998) and Taiwan Island of China ( Chang et al. 2012). The fossil record shows that Rhinopithecus was widely distributed in south, southwest, and central China, and in 2 provinces in northwest China (Gansu and Shaanxi) until about 400 years ago, when increasing human populations accompanied a dramatic range retraction ( Li et al. 2002). Rhinopithecus lantianensis , a largebodied, extinct form, was found north of the Yangtze River (Jablonski and Peng 1993). R. tingianus Mathew and Granger 1923 , reported from Hunan (Wang et al. 1982), is likely a subspecies or antecessor of R. brelichi ( Groves 1970) . Liedigk et al. (2012) note that, because most Rhinopithecus fossils consist only of teeth and jaw fragments, identification to species is challenging.
FORM AND FUNCTION
Form. —The dental formula of Rhinopithecus bieti is i 2/2, c 1/1, p 2/2, m 3/3, total 32 ( Elliot 1913). Rhinopithecus species, together with Semnopithecus , have the largest molars (allo-scaled) among colobines (Pan and Oxnard 2003). Like the proboscis monkey and douc langurs, R. bieti has a distinct presaccus at the forestomach, giving the stomach 4 parts, in contrast to the tripartite stomach of the Presbytini ( Caton 1998) . The vertebral formula is 7 C, 12 T, 7 L, 3 S; the number of caudal vertebrae has not been reported, but presumably is 25 C with some variation as in other Rhinopithecini , e.g., mean 25± SD 0.79, range 23–26 in proboscis monkeys (cf. Schultz 1942; Schultz and Straus 1945).
Craniofacial, dental, and mandibular morphology have been well described and contrasted with other Asian colobines, and with other snub-nosed monkeys (Peng et al. 1984, 1988; Pan et al. 1991, 1995; Pan and Oxnard 2001). Dermatoglyphs (skin patterns on the finger tips, palms, and soles) have been described (Zhang et al. 1981).
Rhinopithecus bieti exhibits neuroanatomical asymmetry in the Sulcus principalis ( Ma et al. 1992), the part of the simian (but not human) brain that connects the dorsoventral and dorsolateral frontal cortex and is thought to be involved in spatial perception. This structure may be involved in postural handedness (see “Function”).
Function. —The foregut of colobines contains symbiotic bacteria that ferment structural carbohydrates: the animal recovers nutrients by digesting the bacteria. Rhinopithecus species consume especially difficult foods high in tannins and other plant toxins (Kay and Davies 1994). In digestion trials of captive R. bieti using plastic markers, mean transit time was 27 ± SD 7.4 h (n = 3) and retention time was 47 ± SD 17 h (n = 3); recently captured individuals, with presumably natural gut flora, had longer transit times and retention times than did long-term captives ( Kirkpatrick et al. 2001). Transit times were longer than reported for other colobines, but retention times were similar (summarized in Kirkpatrick et al. 2001). In captivity, apparent digestibility of dry food consisting of grasses, dicot leaves, wheat–barley biscuits, and lichens ( Bryoria ) was 71–80%, similar to other colobines but below that of ruminant ungulates feeding on lichens ( Kirkpatrick et al. 2001). Fecal analysis accurately determines the relative proportion of leaves and lichens in diets, but not of relative proportions of foods with different digestibilities, such as fruit versus leaves. Lichens and leaves are not nutritional equivalents but both appear necessary for passage through the gut ( Kirkpatrick et al. 2001).
Using restriction fragment length polymorphism analysis, Wu et al. (2010) reported the fecal bacteria of R. bieti included Firmicutes, Proteobacteria, Bacteroidetes, Fibrobacteres, Spirochaetes, and Actinobacteria belonging to 17 genera, in addition to a variety of uncultured and unidentified bacteria. Stomach microflora RNA are enriched in genes for the metabolism of carbohydrates, amino acids, nucleotides, glycans, and vitamins; the metabolism of fatty acids; and many genes involved in cellulose digestion (Zhou et al. 2014). Xu et al. (2016) found a xylanase from R. bieti gut bacteria of the genus Massilia , which may aid salt-tolerance in digestion.
When selecting mature leaves in autumn, R. bieti chooses those with lower fiber content, higher ash, a higher ratio of crude protein to acid detergent fiber, and lower toughness than nonselected leaves; selected and nonselected leaves were not different in fat, crude protein, total phenolics, hemicelluloses, and lignin ( Huang et al. 2010).
Relative to macaques ( Macaca ) and surilis ( Presbytis ), the more prominent symphysial fusion, corpus, and sagittal condylar dimension of R. roxellana and R. bieti produce more efficient biting force on the incisal and posterior canine teeth, with the heavy reaction force born on the temporomandibular joint; the well-developed bizygomatic width and mandibular height suggest that posterior canine function is similarly prominent in both R. bieti and R. roxellan but incisal function is stronger in the former (Pan et al. 1995).
Pan et al. (2011) found only small to moderate proportions of 13 individuals to exhibit hand preference across a variety of tasks. There was little gender difference: although females used the right hand significantly more than males for reaching and carrying, subjects showed overall symmetrical use of hands for this task. All subjects used the right hand significantly more often than equal left- and right-hand use for object manipulation and extractive foraging; they also used the right hand more, but without statistical significance, for extractive foraging, which requires bimanual coordination (Pan et al. 2011). No left-handed individuals were identified in this small sample. The right-handedness for object manipulation in R. bieti was not found for R. roxellana (Zhao et al. 2008 cited by Pan et al. 2011), an interspecies difference that may be related to the greater sexual dimorphism of the former.
In a follow-up study, Pan et al. (2013) compared hand and foot preference in various postures. Among all 11 subjects, right-handedness was stronger in clinging and bipedal postures compared to sitting or quadrupedal postures, although males more often switched from their left hand in the sitting posture to their right hand in the bipedal posture. At the individual level, handedness was not consistent among postures (Pan et al. 2013). Interestingly, more than one-half the test subjects—all genetically related—preferred the left foot for object manipulation, but the small sample size prevented conclusions about this (Pan et al. 2013).
Yu et al. (2011) found evidence in the mitochondrial genome of R. roxellana for positive selection in genes that may help with energy metabolism at high altitude and in cold weather; however, these adaptations were absent in R. bieti , suggesting that the latter had other adaptations that allowed them to exploit even higher elevations and colder weather. Likewise, Wang et al. (2010) found no difference in the evolutionary pattern of the Leptin gene between these 2 high-elevation snub-nosed monkeys, compared to their lowland counterpart, R. avunculus , and other colobines, in contrast to a previous finding of adaptive evolution of Leptin genes in other high-elevation mammals such as the plateau pika ( Ochotona curzoniae ). Leptin, an adipocytederived hormone, is believed to play an important role in energy homeostasis in adaptation to high-altitude environments; its lack of differentiation between lowland and highland colobines suggests that R. bieti has developed other mechanisms to adapt to cold and avoid hypoxia associated with its high-elevation habitats (Wang et al. 2010).
Zhou et al. (2014) identified 105 positively selected genes in the 4 Chinese Rhinopithecus species that were mainly associated with the biosynthesis of fatty acids; signaling pathways for insulin, adipocytokine, and valine, leucine, and isoleucine degradation; and genes for propanoate metabolism, pyruvate metabolism, and lipid binding. These adaptations suggest enhanced energy metabolism to efficiently degrade plant cell wall components (celluloses and hemicelluloses) and absorb and exploit the scarce nutrients typical of the Rhinopithecus diet.
The team also found that more than one-half of the olfactory receptor genes in species of Rhinopithecus were inoperative pseudogenes or gene fragments, a higher proportion than for other primates and mammals; they suggested that the reduction in the olfactory genes in Rhinopithecus might have coincided with the recession of the external nostril, which houses the cilia of olfactory sensory neurons (Zhou et al. 2014).
ONTOGENY AND REPRODUCTION
Ontogeny. —In captivity, sexual maturity occurs at 4–5 years for female Rhinopithecus bieti and 5–6 years for males ( Ji et al. 1998). Immatures ≤ 1 year old have similar gray coat colors and body size, indicating synchronized parturition during March–April; neonates have “brilliant white coats” that begin to have touches of gray by July and are all gray by autumn ( Kirkpatrick et al. 1998:24). Infant time on females, apparently suckling, declined from a peak of 51% of observation time in July to 9% in December.
Locomotion behaviors vary with ontogenic stage (Isler and Grueter 2006 and references therein): arm swinging (semi-brachiation) is seen in juveniles but disappears during adulthood; adults leap more frequently in conjunction with quadrupedal climbing in trees; adults more frequently climb vertical or nearvertical tree trunks of large diameter, which involves consistently reaching with the forelimbs above the level of the shoulder; and young R. bieti sometimes walk bipedally on branches for short distances.
Reproduction. —Copulations occur all year, with peaks in the August–October mating season and a 2nd, nonbreeding peak during the March–April birth season ( Kirkpatrick et al. 1998; Huang et al. 2012; Wang et al. 2012). At Mt. Lasha in the south ( Fig. 3 View Fig ), copulation frequency peaks 1 month after peaks in staple foods, rainfall, and minimum temperatures, and 3–4 months after peaks in high-quality foods ( Huang et al. 2012). At Xiaochangdu, mating begins a little earlier, July–October, also coinciding with peak food availability and temperature at that northern site (Xiang and Sayers 2009). The monthly rate of intermale aggression increases during the mating season but is not significantly correlated with the copulation rate ( Kirkpatrick et al. 1998).
He et al. (2001) measured reproductive hormone secretion during the menstrual cycle and pregnancy of Rhinopithecus bieti by monitoring urinary estrone conjugate (E 1 C), pregnanediol-3-glucuronide (PdG), bioactive follicle-stimulating hormone (FSH), and luteinizing hormone (LH). They found that: (1) E 1 C peaked before ovulation (183.9 ± SE 8.6 ng /mg Cr) followed by (2) an elevation of PdG in the luteal phase (754.4 ± SE 30.6 ng / mg Cr) that was 3–4 times higher than during the corresponding follicular phase (198.3 ± SE 11.4 ng /mg Cr); (3) the peaks of LH and FSH were on the same day, while E 1 C peaked 1–2 days before; (4) FSH levels were higher in the follicular phase than in the luteal phase, when it was only slightly elevated; (5) the mean cycle length was 23.6 ± SE 3.5 days (n = 3) based upon successive urinary LH peaks; (6) based on the interval from the day of the E 1 C peak to the day of parturition, the gestation was 203.7 ± SE 2.5 days (n = 3); and (7) both E 1 C and PdG increased and remained high after pregnancy, with a sharp decrease to basal levels following parturition or midterm abortion. They concluded that, although the pattern of reproductive hormones for R. bieti is similar to that of other Old World monkeys, the hormone concentrations are unique and R. bieti has a longer progestation period ( He et al. 2001).
In captivity, parturition begins in December and peaks in March–May with a median birth date of April 10 ( Cui et al. 2006b). This is similar to the wild, where a 6.5- to 7.0-month gestation results in a birth season centering on March–April at more southerly sites ( Kirkpatrick et al. 1998; Huang et al. 2012) and February–March at Xiaochangdu in the north (Xiang and Sayers 2009).
Interbirth interval in captivity mirrors that in the wild, 428 ± SD 87 days for females that had lost an infant at <1 year of age or experienced a still birth (29% of the sample), and 706 ± SD 71 days for females (71%) whose last infant survived> 1 year, or an overall average of 624 ± SD 150 days ( Cui et al. 2006b). Infant mortality is higher in the wild, 60% ( Kirkpatrick et al. 1998), than in captivity, 30% ( Cui et al. 2006b).
ECOLOGY
Population characteristics. —In 2015 there were 15 known groups (= “bands”; see “Behavior”) in Yunnan, comprising 5 subpopulations of Rhinopithecus bieti that were isolated geographically and genetically distinct across the geographic range of the species ( Li et al. 2015a; see “Population genetics”). These 15 bands did not include the recent observation of a small population in the Yunlong-Tianchi Nature Reserve (see “Distribution”), making a total of 16 extant groups in Yunnan. In Tibet, 3 bands are known, 2 of which are moderately connected and the 3rd isolated (Xiao et al. 2005; Yang et al. 2005).
In a band of about 175 individuals comprising 15–18 onemale units plus a dozen or so unaffiliated individuals at Wuyapuya (Deqing County; Fig. 3 View Fig ), the adult sex ratio was skewed toward females (3.1 females/male); this band had a 1:1 ratio of adults and immatures and a 1:2.5 ratio of yearling juveniles to infants ( Kirkpatrick et al. 1998). The latter ratio suggested a 1st-year mortality rate of 60%, an estimate corroborated by comparing the number of infants estimated for July 1993 with yearlings for June 1994 ( Kirkpatrick et al. 1998).
For a population viability analysis, Xiao et al. (2005) gave group sizes of 13 bands of 30–360 totaling 1,701 individuals with the following estimated population parameters: mortality in year 1 for both female and male infants, 55% ± SD 10.0%; year 2, 15% ± SD 5.0% for both genders; year 3, 10% ± SD 4.0% for both genders; years 4 and 5, 5% ± SD 2.0% for both genders; years 6, 7, and 8 for females 3% ± SD 1.0% and for males 5% ± SD 2.0%; and for older adults 3.0% ± SD 1.0% for both genders. They estimated reproduction rates (= fecundity) of 0.95–1.00 for these 13 bands. For 5 of the bands, they estimated the proportion of males breeding, 28.9% to 48.8%, proportion of males successfully siring offspring, 27.5% to 40.9%, and mean number of litters sired by successful males in an average breeding cycle, 2.2 to 3.2 (Xiao et al. 2005).
Space use. —The ranging pattern of Rhinopithecus bieti has been described as “seminomadic” as bands range over large areas, not remaining long in recently foraged areas and instead seeking new areas ( Kirkpatrick et al. 1998). Most groups have different seasonal distributions or visit patches of known seasonal forage (Zhao et al. 1988; Wu 1993; Yang 2003; Liu et al. 2004; Grueter et al. 2008; Li et al. 2008; Ren et al. 2009). Bands also move daily and seasonally in response to environmental variables and physiographic features such as temperature, slope, and sunlight (Liu et al. 2004; Xiang et al. 2010a; Quan et al. 2011; Huang et al. 2012; Xiang 2013).
In any given year, a band’s home range may be from about 16 km 2 (e.g., Wuyapuya—Kirkpatrick et al. 1998) to 40 km 2 (e.g., Xiaochangdu—Long et al. 1994); but over longer periods a band uses a larger home range, for example, about 100 km 2 over 10 years at Wuyapuya ( Kirkpatrick et al. 1998). Wuyapuya is in Deqing County in the northern part of Baimaxueshan National Nature Reserve and is isolated from the southern portions of the Nature Reserve by Highway 214 and other anthropogenic features.
Grueter et al. (2008 and references therein) summarized home range size of 6 bands that ranged from 9.56 to 25.3 km 2. Across all bands in the Samage Forest where both home range size and group size were known, they were positively correlated ( Grueter et al. 2008). At Samage Forest, by grid occupancy, Grueter et al. (2008) found that a large group of R. bieti (> 400 individuals) ranged over 32 km 2 in a 14.5-month study, and over 56 km 2 in 10 years; home range sizes did not differ during winter, spring, and summer, but were smaller in autumn. Samage Forest comprises the southern part of Baimaxueshan (White Horse Snow Mountain) National Nature Reserve, near the town of Tacheng, and includes the villages of Gehuaqing, Xiangguqing, and Cikatong (Deqing and Weixi counties; Fig. 3 View Fig ). In this part of Baimaxueshan, 5 R. bieti bands are now known ( Li et al. 2015a).
One of the 2 bands at Laojunshan (Mt. Laojun) near Jinsichang (Lijiang and Lanping counties; Fig. 3 View Fig ) also ranged over 32 km 2 (100% minimum convex polygon) in 11 months; mean daily range size was 4.80 ± SD 5.81 ha with a range 0.0145.67 ha (Ren et al. 2009).
There is little evidence of range overlap among bands ( Kirkpatrick et al. 1998) and only 1 band occupies each of the 16 extant band home ranges in Yunnan, as far as is known. However, a comparatively low index of defendability of the Gehuaqing group at Samage Forest suggests that groups are not territorial per se ( Grueter et al. 2008). Grueter et al. (2008) reported that a small band (the “Shikuadi” group) briefly entered the home range of the Gehuaqing band; whether this indicated an overlap of home range or merely a transitory incursion was not clear.
During sleeping and resting, groups arrange themselves into 27–106 m 2 /individual, and disperse over 5,000 –15,000 m 2 /individual while feeding (Wu 1993). Feeding bands arrange themselves into a rough circle with each individual 1-male family unit or all-male unit keeping together and the males aggressively (by glaring or chasing) defending their feeding patch when members of other units approached to within 2–5 m ( Kirkpatrick et al. 1998).
Rhinopithecus bieti chooses tall trees for sleeping, usually on a moderately steep slope with south or southeast sun exposure and protection from wind, avoiding both the creek bottom and the ridge top (Zhao et al. 1988; Liu and Zhao 2004; Cui et al. 2006a). Selection of sleeping sites, and the trees selected, appear to consider both energetic (i.e., temperature, sun and wind exposure) and antipredator potential, the selection pressures being strong enough that R. bieti groups are willing to travel some distance to chosen sleeping sites if suitable ones are not located near foraging areas (Liu and Zhao 2004; Cui et al. 2006a). Otherwise, sleeping sites are at or near foraging areas ( Li et al. 2013). At Nanren (a village in Deqing County north of Wuyapuya; Fig. 3 View Fig ), sleeping sites are usually in the middle of slopes that offer morning sun exposure; in winter they average lower elevation than at other seasons ( Cui et al. 2006a). Sleeping trees are tall emergents (27.5 ± SD 3.2 m) with large diameter at breast height (DBH; 57.9 ± SD 16.9 cm) and broad crown diameters (6.3 ± SD 1.4 m); focal groups choose tall conifers (fir, Abies georgei , or spruce, Picea likiangensis ) over tall oaks, Quercus ; and sleeping sites are changed nightly ( Cui et al. 2006a). At Samage Forest ( Li et al. 2006b), sleeping sites are exclusively in evergreen trees (82% conifers, mainly P. likiangensis and Tsuga dumosa , and 18% evergreen broadleaf trees, Cyclobalanopsis oxyodon and Quercus ); sleeping trees are tall (mean 30.5 ± SD 6.4 m), have high boles (mean 18.4 ± SD 5.2 m), large diameters (mean 62.6 cm ± SD 23.8), and large crown areas (mean 57.9 ± SD 38.0 m 2). Sleeping sites at Mt. Fuhe (Lanping County; Fig. 3 View Fig ) are similar, but the R. bieti there more often uses the same tree on successive nights (Liu and Zhao 2004). R. bieti groups do not relocate after entering sleeping sites, that is, after sunset (Ren et al. 2008).
Mean daily travel distance was about 1.59 km (maximum 2.95 km) over an 8-month period at Wuyapuya, and R. bieti travels farther in warmer months ( Kirkpatrick et al. 1998). At Samage Forest, a band travelled a mean of 1.62 ± SD 0.798 km / day (range 0.578 –4.216 km /day) in a “zigzag pattern with frequent returns to previously visited areas” during a full month follow in September 2006 ( Grueter et al. 2008:1133). Daily path length varies significantly by season. For example, at Samage Forest a band travelled a mean of 985 m per day in winter versus mean 1,721 m (F = 6.426, P = 0.016) in summer ( Grueter et al. 2013a).
At Jinsichang, the band travelled 0.863 –0.962 km /day (based on multipoint fixes of 2 satellite-collared males that were the breeding males of their respective one male unit) that generally represent the cumulative distance from the sleeping site to a morning foraging site, a resting site, and afternoon foraging site, and to the next sleeping site (Ren et al. 2008).
Rhinopithecus bieti bands usually travel in single-file to sleeping and feeding sites, avoiding open areas, often following known trails in a regular sequence, according to seasonal phenology of food resources (Wu et al. 1988; Wu 1993). Bands spend several hours at a foraging area before travelling to another, or to a sleeping site, so that by satellite telemetry, foraging and sleeping sites appear as clusters of point locations (Ren et al. 2008).
Rhinopithecus bieti is 1 of the highest ranging and most northerly primates , occupying elevations from 2,625 to 4,600 m (n = 5 groups—Kirkpatrick and Grueter 2010) with occasional forays up to 4,700 m, far above the tree line (at Wuyapuya— Long et al. 1994). By contrast, R. strykeri has been found at 2,400 –3,300 m in Myanmar ( Chen et al. 2015) and 2,600 –3,100 m in mainland China ( Ma et al. 2014), while R. roxellana lives at 1,000 –4,100 m (Kirkpatrick and Grueter 2010). In Tibet, a large group (N = 174) of R. bieti has been filmed walking over a mountain pass through steep, rocky terrain above 4,700 m between Nanren and Wuyapiya to reach forest patches on the other side (Lian-Xian, personal communication, September 2018).
At Xiaochangdu in the north, day length ranges from 12.2 to 16.0 h and monthly mean temperature ranges from 12.5°C in August to −3.6°C in January; annual maximum and minimum temperatures are 26.9°C and −15.4°C, respectively (Xiang et al. 2010a). Snow is present for 4–6 months of winter throughout the range of R. bieti , but is deeper and lasts longer in the north, although wind and sun exposure on south-facing slopes often reduce snow cover along ridge tops (Xiao et al. 2003). In contrast, at Mt. Longma in the south, the average annual temperature is 8.1°C and monthly average temperatures range from 14.3°C in August to 2.4°C in January (Li and Yang 2009).
Xiao et al. (2003) classified R. bieti range into 3 broad types: (1) dark-coniferous forests of fir, A. georgei , and spruce, P. likiangensi s; (2) mixed coniferous and broadleaf forests; and (3) Yunnan pine ( Pinus yunnanensis ) forest. R. bieti avoid the pine forests, leaving the fir-spruce and mixed coniferous–broadleaf types as suitable habitat. Other authors classify the coniferous forest band as fir-larch, Larix potaninii (e.g., Long et al. 1994).
Rhinopithecus bieti mainly uses coniferous or mixed conif-erous–broad-leaved forest above the broad-leaved forest and agricultural zones and below tree line ( Fig. 4 View Fig ); these zones increase in elevation from south to north, defining potential habitat. At Mt. Longma, the fir-larch band is 3,000 –3,500 m, while at Xiaochangdu it is at 3,700 –4,200 m ( Long et al. 1994). The Jinsichang R.bieti band ranges between 3,200 and 3,630 m (Yang 2003) and the Wuyapuya band 3,500 –4,600 m ( Kirkpatrick et al. 1998). At Samage Forest, the Gehuaqing band ( Fig. 3 View Fig ) ranges over 2,600 to 4,000 m and uses lower slopes in spring to access new leaf growth ( Li et al. 2008). For all subpopulations, the elevational range is generally restricted by human disturbance and altered habitats at the lower elevations, and by tree line at the upper elevations ( Long et al. 1994). However, moderate human activity, such as herding livestock in alpine meadows, does not dissuade R. bieti from using adjacent forest edge habitats; nor does absence of human activity in the forested zone, as in winter when livestock have been brought to valley bottoms, induce R. bieti to use lower-elevation forest habitats ( Kirkpatrick et al. 1998; Xiang et al. 2007b; Zhong et al. 2008). In some northerly populations, bands remain at high elevation through the winter, whether for access to food on slopes and ridges where wind reduces snow depth ( Grueter et al. 2012), or for the warmth of sunlight on high, south-facing slopes (Quan et al. 2011), as noted above.
Rhinopithecus bieti habitat quality ranges from more productive in the south to marginal in the north, which is “climatologically harsher than any other inhabited by non-human primates ” and where floral diversity is lower (Xiang et al. 2007b:1141). For example, Long et al. (1994:246) noted that floristic diversity was greater in the southern end of R. bieti range, with 24 species of trees and tree-like shrubs at Mt. Longma, compared to 5 “core” species at Xiaochangdu. In more northerly populations, at all seasons, R. bieti fecal density is correlated with the abundance of arboreal lichens, mainly of the genus Bryoria , a staple food item especially in winter (Zhong et al. 2008).
Rhinopithecus bieti habitat at Xiaochangdu comprises 4 habitat types: (1) primary conifer forest (mainly P. likiangensis and Abies squamata trees with a thin understory of bushes <2 m in height), (2) secondary conifer forest (as above but selectively logged with a denser understory), (3) larch forest (mainly L. griffithiana and Rhododendron trees with a dense shrub layer of mostly Rhododendron usually> 2 m), and (4) evergreen broadleaf forest (mainly Quercus aquifolioides trees) and low density shrubs usually ≤ 2 m high (Xiang et al. 2007b).
At Wuyapuya, R. bieti habitat is characterized by fir, A. georgei , and oak, Q. pannosa ( Kirkpatrick et al. 1998) . R. bieti habitat in a mixed coniferous–broadleaf forest in the Samage Forest includes 80 tree species (23 families), the 3 dominant tree species by basal area being A. georgei , Cyclobalanopsis cf. gambleana , and P. likiangensis ; 1 of the conifers ( Larix ) and 73% of the broadleaf trees are deciduous (Grueter et al. 2009). The Samage Forest habitat of R. bieti has 5 main forest types ( Li et al. 2008): (1) mixed deciduous broadleaf and conifers, (2) pinedominated ( P. yunnansis ), (3) evergreen broadleaf dominated by Cyclobalanopsis spp. in the canopy, (4) cool temperate fir-dominated ( A. georgei ), and (5) montane sclerophylus oak-dominated ( Q. pannosa ). Of these, they preferred the mixed deciduous–coni-fer type and usually avoided the others ( Li et al. 2008).
A site at Xiangguqing (east of Tacheng in Fig. 3 View Fig ) illustrates the habitat diversity at a mesoscale: this Samage Forest site has essentially 2 habitat types: (1) a small bamboo forest ( Fargesia yunnanensis ) with T. dumosa and Rhododendron yunnanensis the dominant tree species, and (2) a more extensive fir ( T. dumosa )–rhododendron ( R. rubiginosum ) forest with a shrub layer dominated by Lyonia ovalifolia and Enkianthus deflexus (Ren et al. 2012a) .
In autumn, members of this population choose slopes of 20° to 40° at elevations of 2,700 –3,300 m with a tree canopy cover of 40–80% in deciduous broadleaf or mixed deciduous broadleafconiferous forest with DBH> 20 cm; within this habitat, they use locations> 500 m from human disturbance ( Li et al. 2006a). Jinsichang habitat occupies a nearly continuous primary coniferous forest belt (3,200 –4,100 m) dominated by fir ( A. georgei ) and spruce ( P. likiangensis ) below 3,700 m and nearly pure fir forest above that level ( Long et al. 1994). R. bieti habitat at Mt. Longma is dominated by fir ( A. georgei ), larch ( L. potaninii ), oak ( Q. pannosa ), and rhododendron trees with a wide variety of deciduous trees and tree-like shrubs ( Long et al. 1994).
Diet. —As a broad generality, Rhinopithecus bieti relies heavily on immature leaves of monocots and dicots in spring, fruit in summer and autumn, and lichens ( Fig. 5 View Fig ) in winter (Grueter et al. 2009). Wu (1991) characterized the diet of captive and wild R. bieti as (1) leaves and stems of gramminoids, (2) hyphae of lichens, and (3) buds, bark, small stems, and fruits of angiosperm trees, but not conifer needles. R. bieti selects both mature and young leaves that have higher crude protein and lower fiber (neutral detergent fiber, acid detergent fiber, and celluloses) and tannin than nonfood items (Li and Yang 2009; Huang et al. 2010).
Dietary breadth decreases with increasing latitude and altitude (reviewed by Grueter et al. 2009): at Xiaochangdu, R. bieti feeds on 25 vascular plant species, ≥ 3 species of lichens (a staple dietary constituent throughout the year, although it prefers vascular plant foliage) and its diet includes at least 7 species of invertebrates (Xiang et al. 2007b). There, in contrast to more southerly populations, lichens account for 47–64% of the diet (Yang et al. 2005). At Wuyapuya, its diet includes 20 species of plants ( Kirkpatrick 1996). In the Samage Forest, although R. bieti feeds on 150 different plant parts from at least 94 species and 38 families, 6 genera ( Acanthopanax , Sorbus , Acer , Fargesia , Pterocarya , and Cornus ) provide most of the diet; it highly selects a few uncommon angiosperm tree species (Grueter et al. 2009). R. bieti eats bamboo shoots in the summer and bamboo leaves ( Fargesia ) throughout the year, forages mushrooms, and digs up tubers; 1 consumed the flesh of an unidentified flying squirrel ( Sciuridae ) and 1 ate a bird’s eggs (Grueter et al. 2009). Zhao et al. (2009) extended the list to include 2 lichen species and 110 vascular plant species. At Xiangguqing (east of Tacheng; Fig. 3 View Fig ), in a 2-month (June–July) diet study and provisioning experiment, R. bieti ate 32 wild plant food species, with bamboo shoots, Fargesia , especially prominent and preferred over lichen, Usnea longissima (Ren et al. 2012a) .
At Jinsichang (Yang and Zhao 2001), diet includes, in order of frequency: (1) bamboo leaves, (2) dicotyledon leaves, (3) lichens, including fruticose ( Bryoria , U. longissima ) and foliose (family Stictaceae ) lichens, (4) seeds, mainly from Acanthopanax evodiaefolius , (5) other, including conifer needles, probably taken accidentally, and a rodent (Yulongxuen red-backed vole, Eothenomys proditor ). At Mt. Longma, R. bieti consumed ≥ 97 plant species (based on indirect observations such as trail signs—Huo 2005). This group’s winter diet includes 45 plant species of 26 families: Fargesia papyrifera , 19.8%, Qiongzhuea rigidula , 4.9%, and Fargesia pallens , 4.5%, while main lichens were U. longissima , 10.4% and Usnea florida , 8.4%, by frequency in scat analysis (Li and Yang 2009).
Diseases and parasites. —Of 8 Rhinopithecus bieti captured in the wild and sent to Kunming zoo in 1987–1992 ( Ji et al. 1998), all tested negative for Simian Acquired Immunodeficiency Syndrome Retrovirus ( SRV), Herpesvirus B, and tuberculosis (TB) and all were free of pathogenic bacilli. A Trichocephalid parasite was found in their feces (see below). Blood profiles (red and white corpuscles, platelets, hemoglobin, etc.) were similar to those of other primates . Values for urea nitrogen, creatine, glutamine transaminase, glutamyltranspeptidase, lactate dehydrogenase, and alkaline phosphatase levels were higher than those of humans and rhesus macaques ( Macaca mulatta ). Glucose levels were considerably higher than those reported for humans and M. mulatta . Uric acid levels and triglycerides were lower than those recognized for humans and rhesus macaques. The values of lactic dehydrogenase (LDH), glutamate oxaloacetate transanunase (GOT), and glutamate pyruvate transaminase (GPT) differed from those of other monkeys and humans ( Ji et al. 1998).
Lin et al. (2016) mentioned heavy nematode loads in R. bieti . In 1 study, 11 of 16 (68.75%) captive R. bieti had Trichuris eggs, but no other gastrointestinal parasites were identified ( Li et al. 2015b). However, wild R. bieti have higher parasite loads: Trichuris egg densities are different between adults and juveniles, infections of Ancylostoma and Oesophagostomum vary seasonally (highest infection rate is in the raining season); and Capillaria hepatica and Ascaris are occasionally observed ( Li et al. 2014b).
Hou and Peng (1989) named a new, apparently host-specific species of Trichocephalus (Trichocephalidae) found in wild R. bieti from Weixi County (near Tacheng in Fig. 3 View Fig ): T. rhinopithecus . The 1st flea reported on R. bieti , apparently a host-specific subspecies, was named Vermipsylla parallela rhinopitheca ( Li 1985) . Using different sleeping sites on consecutive days may avoid parasite transmission by preventing the accumulation of feces ( Cui et al. 2006a; Li et al. 2013).
Interspecific interactions. —Rhesus macaques are sympatric with Rhinopithecus bieti in the Samage Forest. Since at least 14 of about 22 plant species eaten by the rhesus macaque are also eaten by R. bieti , both species prefer fruits, and both use mixed broadleaf deciduous–conifer forest most frequently, there is potential for niche overlap ( Grueter et al. 2010). However, they use space and some resources differently: (1) they maintain spatial separation (mean elevation of R. bieti locations was 3,218 ± SD 188 m versus 2,995 ± SD 215 m for the rhesus macaque and the latter was sometimes seen in meadows while R. bieti was not), (2) the rhesus macaque often uses evergreen broadleaf ( Cyclobalanopsis -dominated) forest while R. bieti rarely does, (3) the rhesus macaque raids crops and R. bieti does not, (4) groups of the 2 species usually kept space between them (mean 2.4 ± SD 2.5 km), and (5) when a close encounter did occur, a confrontation resulted in the rhesus macaques moving away. The authors suggested that, “the coexistence of the two taxa is facilitated via differential macrohabitat use and spatial avoidance” ( Grueter et al. 2010:516).
Eagles ( Aquila ) and buzzards ( Buteo ) are reported to prey on R. bieti ( Bai et al. 1987; Liu and Zhao 2004; Li et al. 2013). Cui (2003:52) described a hawk ( Buteo ) attack on a 1-male group, which reveals the reactions and sex-age-specific roles of R. bieti individuals to this predator: “On seeing a buzzard, Buteo sp., flying from a distance, adult monkeys in the group instantly emitted alarm calls and the group members immediately dropped from the upper to the middle/lower canopy of the fir trees they were in. When the buzzard was soaring over the canopy in search of them, the monkeys continued to descend and emitted loud alarm calls ….”
The hawk continued alternately perching and flying around the group as they progressed lower in the trees. After 13 min the hawk attacked, upon which, “...the adult females enfolded the infants in their arms, the juveniles clasped their mothers tightly and all the monkeys emitted strident calls. Meanwhile, the adult male attacked the buzzard around the fir tree, opening his mouth and showing his large, sharp canines and giving loud calls.” The hawk finally flew away after a total encounter time of 14 min.
Leopards ( Panthera pardus ), if they still occur in R. bieti range, probably prey (or preyed) on them, as they have on R. roxellana (see discussion in Cui et al. 2006a). Liu and Zhao (2004:246), noting the apparent absence of leopards at Mt. Fuhe ( Fig. 3 View Fig ), speculated that antipredator behavior around sleeping sites (see above) “may be at least partly viewed as a trace of cultural tradition [i.e., instinct] ...developed in the past under heavy predation pressure.”
BEHAVIOR
Grouping behavior. — Rhinopithecus bieti has a 2-tiered grouping system in which from 11 to 26 “family” groups (or 1-male units) of 3–17 individuals including several mature females, and usually 1 or more all-male units, form variable groups (“bands”) of ≤ 480 individuals ( Kirkpatrick et al. 1998; Xiao et al. 2005; Cui et al. 2008; Grueter 2009; Ren et al. 2012a; Wang et al. 2012; Li et al. 2014a). Bachelors within the band are threats to the males of 1-male units, and the frequency of male aggression is elevated when bachelors are present (Grueter 2009).
Ren et al. (2012a) observed 12 fission–fusion events during 5 years of study of the band at Xiangguqing (Samage Forest), which had grown to 480 individuals by 2008; 11 of these events also involved fission and subsequent fusion of the all-male unit, with transfer of males between subbands. All splits were during June–July and occurred in a forest of bamboo, a preferred food. June–July is after the birthing season when lactating females have high energy demands. In each case, the band split into 2 subbands of different but not greatly disparate sizes that moved independently for 2–11 days. No aggression or other unusual behaviors were seen at the time of the splits, and predators or anti-predator behavior were not observed. The investigators concluded that the fissions allowed more efficient exploitation of a limited food source, while fusions allowed more band-level social cohesion when food sources were more evenly distributed (Ren et al. 2012a). Nevertheless, 1-male units defend feeding patches from other units within a band; observed depletion of feeding patches suggests the potential for competition for food resources and may imply a food-based limitation on carrying capacity (Grueter et al. 2009).
Reproductive behavior. —Females solicit copulation by “prostration plus glancing laterally” or “sitting plus head moving up and down” displays, and males solicit by a “grunt bared-teeth display” (Cui and Xiao 2004). Ejaculatory copulations are rare (e.g., mount to ejaculation ratio 5.2 ± SE 4.0 in captivity—Cui and Xiao 2004) and seem to fit the “multiplemount ejaculation” copulatory pattern; only those during the summer–autumn mating season appear to result in pregnancy (Cui and Xiao 2004; Wang et al. 2012).
Although colobine females usually give birth at night, Ding et al. (2013:5) witnessed a primiparous female parturition in the daytime: the parturition stage lasted 4.5 min, after which the female “skillfully severed the umbilical cord, ingested the placenta, and held and licked the newborn infant.” A multiparous female assisted in the delivery, watched with “great interest” by juvenile females in the 1-male unit ( Ding et al. 2013).
Communication. — Rhinopithecus bieti can be very vocal, but groups are usually relatively quiet, communicating mainly by sight using eye contact, positions, and gestures; while sleeping and foraging, “their murmurs can be only heard within a short distance such as 40–80 m ” (Wu et al. 1988:383). Males use a distinct, loud bray during intense internal aggression and other stressful situations ( Kirkpatrick et al. 1998; Grueter 2003). When entering sleeping sites, R. bieti is even quieter than at other times and maintains silence during the night, presumably an antipredator strategy (Liu and Zhao 2004; Cui et al. 2006a).
Miscellaneous behavior. — Rhinopithecus bieti is moderately terrestrial, spending 20% ( Grueter et al. 2013b) to 35% ( Kirkpatrick 1996) of time on the ground; most terrestrial locomotion is associated with travel ( Grueter et al. 2013b). Combining 2 studies in the Tacheng area, as a daily average during daylight hours, R. bieti spends 35–39% feeding, 29–33% resting, 15–19% moving, and 13–14% in social activities (Ding and Zhao 2004; Grueter et al. 2013a). Li et al. (2014a) provided the following time allocation by sex and age class at Xiangguqing in the Samage Forest: adult females spend more time feeding (44.8%) than adult males (39.5%), juveniles (39.1%), and infants (14.2%), and allocate significantly more time to feeding than to any other activity; conversely, adult males allocate more time to miscellaneous activities (12.5%) than do adult females (3.8%).
At the higher, northernmost site, Xiaochangdu ( Fig. 3 View Fig ), where dietary choice is more restricted, R. bieti allocates more time to feeding (33% to 59% of daylight, by month) and foraging (feeding time + travelling time: 62–81% by month). Foraging time is, however, independent of environmental variables (e.g., temperature, day length, and food availability, which are corelated), while nonforaging time (resting + “other”) responds to these seasonal changes (Xiang et al. 2010a). This shows that, even in the far north of its range, R. bieti finds sufficient forage in about 10 h/day regardless of season, allowing it to spend more time in resting and social behaviors as day length increases.
Conversely, at one of the southernmost sites, Mt. Fuhe (Liu et al. 2004), although R. bieti spends similar amounts of time travelling (15% ± SD 2.8% of daylight) and “other” activities including playing and grooming (16% ± SD 2.5%), it spends less time feeding (30% ± SD 4.8%) and more time resting (41% ± SD 3.2%).
Grooming ( Fig. 6 View Fig ) occupies 8.4% of observational time for adults and 6.1% for all age-sex classes; females both groom and are groomed more than males and they spend more time grooming and in close proximity to others than males. Conversely, juveniles groom and are groomed less than adults ( Kirkpatrick et al. 1998; Grueter 2009). The “high level of grooming, primarily among females, suggests strong female-female relationships” (Kirkpatrick and Grueter 2010:107). As well, the high amount of grooming activity involving males compared with males of other colobines suggest a higher level of affiliation with females (Kirkpatrick 1998; Kirkpatrick and Grueter 2010). Infants are groomed 6.9% of the time, the highest proportion among all agesex classes ( Li et al. 2014a).
In winter, R. bieti allocates more time to feeding on the poorer diet, and less to resting ( Grueter et al. 2013a). Social play takes up 3% of infant time in May and peaks at 24% in August ( Kirkpatrick et al. 1998). Huddling together is common but only night length ( Li et al. 2010). The adult male of each 1-male unit sleeps alone; however, female–juvenile and female–infant dyads usually huddle during the night, and group huddling (up to 8 individuals) increases with decreasing temperatures during winter ( Li et al. 2010).
occupies only about 4% of observation time ( Kirkpatrick et al. 1998).
Adult and juvenile females residing in the same 1-male unit provide allomaternal care by transporting, grooming, and holding infants ( Kirkpatrick et al. 1998). Allonursing is rare but has been observed (Ren et al. 2012b). Adult males rarely groom infants or juveniles ( Kirkpatrick et al. 1998), although they passively tolerate infants in close contact, as in Fig. 1 View Fig . Male allocare, ranging from carrying to guarding infants, peaks during the early spring, an energetically stressful season when food is scarce and parturient females are lactating, and ebbs in late summer, when female allocare increases (Xiang et al. 2010b). Xiang et al. (2010b) interpret the males’ allocare and the females’ willingness to relinquish their infants to male care, as an adaptive response to a highly seasonal, high-elevation environment that promotes survival of infants and mothers.
Xiang and Grueter (2007) observed a case of adult male infanticide and cannibalism on a 1-month-old infant, but lack of information on parentage prevented conclusions as to whether this isolated incident fit within the sexual selection hypothesis (cf. Hrdy 1979) or was simply a case of social pathology or accident. Females may carry infants that have died for days if they died when 1 month old but only briefly (e.g., 1 day) in the case of a stillborn infant, and not at all in the case of an aborted fetus ( Li et al. 2012).
In captivity, agonistic interactions occurred infrequently (mean rate of 0.30 per hour), of which 65% involved dyads and 35% involved 3 or more individuals ( Grueter 2004). Most (72%) of aggressive events (threat, chase, contact) were unidirectional and the rest bidirectional; afterwards, post-conflict reconciliation was the norm (i.e., 71% of pairs reconciled and 9.5% were dispersed—Grueter 2004).
At night, at all seasons, it takes about 20 min for members of a group to fall asleep after they enter a sleeping site, and the annual mean sleeping time, 11.5 h (± SD 1.3), is correlated with
GENETICS
Cytogenetics. —The diploid number in Rhinopithecus is 2n = 44 ( Bigoni et al. 2004), as with most other colobines (e.g., Colobus , Trachypithecus , Presbytis , Semnopithecus , and Pygathrix , but not Nasalis : 2n = 48—cf. Harding 2015).
Molecular genetics. —Mirroring morphology, Rhinopithecus bieti occupies a genetic monophyletic clade of oddnosed monkeys, or Rhinopithecini , that also includes the proboscis monkey, simakobu, and douc langurs (Sterner et al. 2006; Whittaker et al. 2006). Zhou et al. (2014) found that a duplication of genes for pancreatic RNase (encoded by RNASE1) in colobines (needed to digest and recover nutrients from the high concentrations of bacterial RNA derived from the symbiotic microflora in the stomach) occurred in 1 mutation event in the ancestor of extant colobines; however, they also found RNASE1 pseudogenes in all studied Asian colobines that were not present in African colobines, and inferred that the retrotransposition event occurred after the African-Asian colobine split, but before the divergence of extant Asian genera.
Based on mtDNA, Meyer et al. (2011) dated the divergence of the Rhinopithecus + Semnopithecus + Nasalis clade from Trachypithecus and Presbytis at about 12.3 million years ago, and Nasalis + Rhinopithecus from Semnopithecus about 10.2 million years ago. Based on mitochondrial and nuclear DNA, Rhinopithecus diverged from Nasalis + Simias + Pygathrix about 7.3 million years ago, R. bieti and R. strykeri form a sister clade to R. roxellana and R. brelichi and these 4 form a sister clade to R. avunculus , which is basal ( Liedigk et al. 2012; Yang et al. 2012). Incongruence between mitochondrial and nuclear DNA suggests ancestral introgression among several of these taxa, including R. bieti + R. strykeri and R. avunculus ( Liedigk et al. 2012) . Various genetic analyses have, however, reached different conclusions regarding relationships within Rhinopithecus (reviewed in Wang et al. 2013).
More recent whole-genome sequencing of the 4 Chinese Rhinopithecus species showed that their genetic diversity was much lower than for all other reported nonhuman primates (heterozygosity 0.025–0.07% in Rhinopithecus versus 0.085–0.24% in other nonhuman primates ): R. brelichi > R. roxellana > R. bieti > R. strykeri (Zhou et al. 2014) . Zhou et al. (2014) produced a maximum likelihood phylogeny supporting a separation between the 2 northeastern species, R. roxellana and R. brelichi , and the 2 Himalayan species, R. bieti and R. strykeri ( R. avunculus was not included in the analysis). They estimated that the northeastern and Himalayan species diverged 1.60 million years ago, coinciding with the Yuanmu movement of the Tibetan plateau uplift, followed by the split of the 2 Himalayan species beginning about 0.3 million years ago, possibly associated with the cold climate of the penultimate Pleistocene glaciation (Zhou et al. 2014).
Population genetics. — Rhinopithecus bieti exhibits adequate haplotype diversity (H) and nucleotide diversity (π) in mtDNA (H: 0.948 ± SD 0.006; π: 0.036 ± SD 0.018), similar to R. roxellana (H: 0.845 ± SD 0.026; π: 0.0 34 ± SD 0.017) and both have higher diversity than R. brelichi (H: 0.457 ± SD 0.084; π: 0.014 ± SD 0.007) in which genetic diversity is low enough to be of conservation concern (Yang et al. 2012).
In molecular and mtDNA from 11 of the extant population fragments (not including the recently rediscovered Mt. Wubao group; see “Distribution”), R. bieti resolves into 2 main clades: the most southerly Fuhe–Lasha–Longma groups comprise the most genetically distinct and isolated clade, while the others comprise 4 distinct subclusters that show decreasing genetic isolation-by-distance from south to north, which reflects anthropogenic habitat barriers ( Liu et al. 2007, 2009; Li et al. 2015a). Liu et al. (2007) hypothesize that the genetic history of R. bieti includes an initial divergence between the 2 main clades 1.0–0.7 million years ago (possibly associated with the Late Cenozoic uplift of the Tibetan Plateau), secondary contact after this divergence as a result of a population expansion 0.16–0.05 million years ago, and population reduction and habitat fragmentation in the very recent past. Genetic differentiation (F ST) among the 5 subpopulations was significant, ranging from 0.1104 to 0.6597, and the migration rate between each subpopulation was low, ranging from 0.0028 to 0.0092, giving additional support to genetic isolation. Although genetic profiles of several individuals suggested potential admixture among subpopulations, Liu et al. (2009) did not find any 1st-generation migrants. There was no genetic evidence of recent population declines, and divergence time estimates ranged from 950 to 21,550 years among subpopulation pairs ( Liu et al. 2009).
CONSERVATION
The Rhinopithecus species of mainland China occurred across south, southwest, and central China, and in 2 provinces in northwest China (Gansu and Shaanxi) until about 400 years ago, but suffered dramatic range contractions because of: (1) the rapidly increasing human population, especially during the 20th century; (2) wars in the 1st one-half of the 20th century; (3) deteriorating environments and accelerated deforestation; and (4) hunting monkeys for food, medicine, and income ( Li et al. 2002).
Rhinopithecus bieti was believed extinct because no observations were reported after the original description (MilneEdwards 1897) until 1960 when 8 skins were obtained from hunters in Deqin County, Yunnan (Peng et al. 1962). The national government established 2 protected areas for R. bieti : the Hongla Xue Shan Mountain (Hongla Snow Mountain) National Nature Reserve in Tibet and Baimaxueshan (White Horse Snow Mountain) National Nature Reserve in Yunnan. Baimaxueshan Nature Reserve, established in 1983, was expanded in 2002 to encompass the Samage Forest ( Fig. 3 View Fig ). The Yunlong and Tianchi nature reserves in the south are provincial. In Tibet, at least 3 subpopulations occur within the Hongla Xue Shan National Nature Reserve, established in 1993 (Xiang et al. 2007a). These 4 nature reserves are units of the Three Parallel Rivers of Yunnan World Heritage Site as designated by the United Nations Educational, Scientific and Cultural Organization (United Nations Educational, Scientific and Cultural Organization 2003).
About 3,082 km 2 or 18.86% of the total landscape in the current distribution area (not including the recently discovered group at Mt. Wubao) of R. bieti is suitable habitat (Wang et al. 2011). This includes Cangshan (Cang Mountains), with 19 peaks> 3,500 m, that Long et al. (1994) found to be suitable habitat, although currently unoccupied, as noted above (see “Distribution”). Based on GIS analysis of present and historical distribution, bioclimatic zones, human influence, topography, tree cover, and protected-areas distribution, Nüchel et al. (2018) showed a vastly larger area of potential habitat to the east and west (i.e., beyond both the Chanjiang [Yangtze] and Lancangjiang [Mekong] Rivers). Several nature reserves already exist in this area, including the large Haba Xue Shan (Haba Snow Mountain) Provincial Nature Reserve that spans Yangtze River on both sides of the Tiger-leaping Gorge, and Pudacuo National Park. These reserves, together with the Cang Mountains to the south, may offer space for population expansion without undue social and economic hardship, if a translocation project were to be considered.
The overall population trajectory since rediscovery in 1960 was in decline until protections became effective in the 1980s, when some subpopulations began to increase, while others continued to decline or became extirpated ( Bai et al. 1987; Long et al. 1994; Xiao et al. 2003, 2005; Yang et al. 2005; Li et al. 2009; Liu et al. 2009; Zhang et al. 2016). Spatially limited, short-term surveys began in 1979, but the 1st comprehensive survey in 1987– 1992 revealed 20 bands or subpopulations ( Long et al. 1994). Surveys completed in 2000 found no sign of monkeys in several areas where they had previously existed, but 3 new bands were found, leaving 11 extant, of which at least 5 were isolated by barriers such as low-elevation farmland, roads, and higher-elevation grazing lands (Xiao et al. 2003). The total population of R. bieti recovered from 800–900 in 1986 ( Bai et al. 1988) to about 1,700 individuals in 13 bands by 2002 ( Ding et al. 2003) and 2,500 based on surveys during 2004–2009 (Wong et al. 2013 citing Y.-C. Long, unpublished data). In 2012, the population in Tibet and Yunnan totaled> 3,000 (Yong-Cheng Long interviewed in Platt 2013), and about 3,500 in early 2016 (Yong-Cheng Long cited in DWĦffi 2016). This suggests a gradual, sustained rate of increase (λ, lambda) of 1.05 over the last 3 decades.
The International Union for Conservation of Nature and Natural Resources lists R. bieti as “Endangered” (Bleisch and Richardson 2008). The Convention on International Trade in Endangered Species of Wild Fauna and Flora (2014) includes R. bieti on Appendix I.
Under Chinese law, R. bieti was designated in the “first class of protected animal” category in 1977. In Yunnan, human activities encroaching on or within R. bieti habitat, such as hunting and logging—formerly with official approval even within protected areas—are the major threats ( Long et al. 1994; Xiao et al. 2003, 2005; Xiang et al. 2007a). Li et al. (2009:477) remarked that, “All the local mountainous residents within the range of the Yunnan snub-nosed monkey (including Yi, Lisu, Tibetan, Pumi, Naxi, Bai, Molimosuo, and Han) have a long hunting history. Although the government’s Wildlife Protection Law was enacted in 1987 [which came into effect in 1989], the law was not fully enforced, and poaching is a key threat...” Li et al. (2009) also mentioned the high density of snares encountered in every survey. National and provincial governments encourage the maintenance of cultural traditions by ethnic minorities ( Goodman 2009) but, as in many parts of the world, this social goal is in conflict with conservation of endangered species.
Threats include use of pesticides: Long et al. (1994) gave an example of forest spraying to control a moth larvae infestation, after which 5 monkey carcasses were found in the Yiyong area, where monkeys have never been seen since. Threats in Tibet include habitat destruction through wood extraction, uncontrolled forest and grass fires, cutting of oak ( Quercus aquifolioides ) branches, “inappropriate attitudes to R. bieti when people carry out economic activities in the forest” and hunting (Xiang et al. 2007a:525). For example, during 347 days of survey from 2002 to 2005, steel snares that could capture R. bieti (though not necessarily set for them) were found on every survey at Milaka; conversely, at Zhina and Xiaochangdu, steel snares were never seen (Xiang et al. 2007a). The difference appeared to be that the last 2 sites were near and within a sacred mountain (Damyon Garpo), respectively, and the 1st was not (although all are within a national nature reserve). Mountains may be designated as sacred by a “living Buddha” or Rinpoche, within limits prescribed by tradition and according to wishes of a local council.
Many of the resources used by R. bieti are also collected by local people, including timber for house-building and fuel wood and plants for Chinese traditional medicine and food, suggesting a need for management to avoid conflicts (Zhao et al. 2009). Yu et al. (2015), identifying 166 medicinal plant species that Tibetan people collect within the Hongla Xue Shan National Nature Reserve, proposed management strategies to avoid conflict with R.bieti while allowing this culturally and economically important activity. Nevertheless, although R. bieti avoids areas of intensive human activity, they do accommodate to modest human presence when not disturbed, as at Tacheng ( Figs. 1 View Fig , 5 View Fig , and 6).
Xiang et al. (2007a) warned of loss of gene flow by mentioning that National Highway G214, Milaka village and farmland form a barrier to dispersal between Tibet andYunnan populations ( Fig. 3 View Fig ). Presence of feces shows that R. bieti move between Xiaochangdu and Milaka populations and it was recommended that this corridor should be protected (Xiang et al. 2007a). The Jinsichang population ( Fig. 3 View Fig ) is isolated and the groups at Mt. Fuhe, Mt. Longma, and Mt. Wubao are isolated from other subpopulations and may be isolated from each other by low-elevation barriers (Xiao et al. 2003). Restoration of habitat along corridors between the Jinsichang and Mt. Fuhe–Mt. Longma population clusters and between the latter and the Samage Forest cluster, to improve connectivity for gene flow has been proposed ( Li et al. 2015a).
In Yunnan, the area of suitable habitat for R. bieti decreased from 4,169 km 2 in 1958 to 1,887 km 2 in 1997, a 31% decline, mainly because of logging and forest clearing to create highelevation summer grazing lands (Xiao et al. 2003). Xiao et al. (2005) calculated higher than 50% probabilities of extinction within 82 years for 5 of 13 extant bands.
On the other hand, sufficient suitable habitat remains for a population of ≤ 5,250 if all bands’ territories in Yunnan and Tibet were filled to their assumed carrying capacities calculated by Xiao et al. (2005:table 3). Given that some 20 bands have been known recently (see “Distribution,” above), the potential exists for population growth if protection from poaching is provided and if habitat loss is limited. There may also be possibilities of (1) expansion to the north within the historic range and (2) reintroduction to the Cang Mountains west of Dali (see “Distribution,” above).

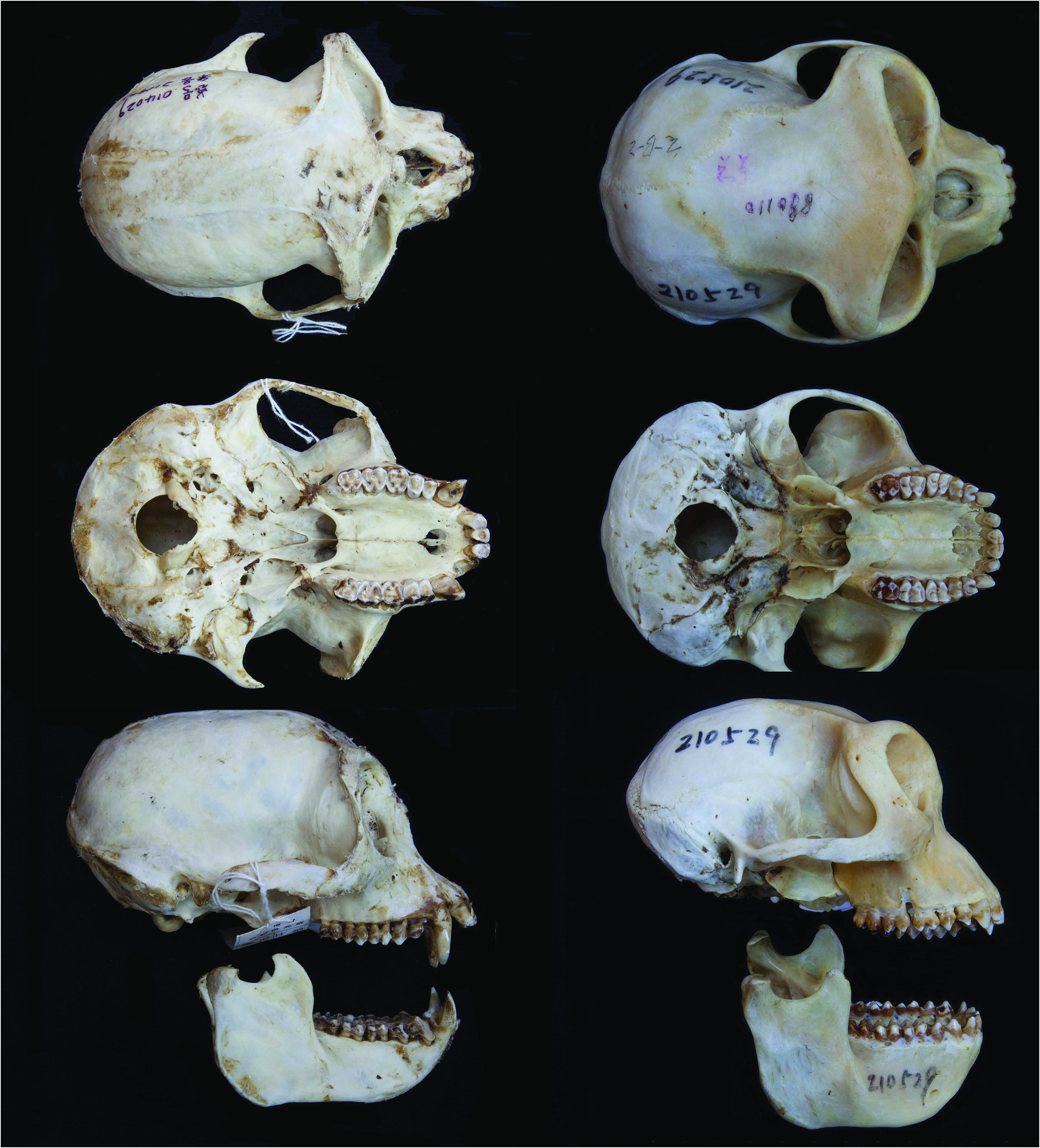




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Rhinopithecus bieti Milne-Edwards, 1897
| Harding, Lee E. & Han, Lian-Xian 2018 |
Rhinopithecus (Rhinopithecus) bieti
| : Jablonski and Peng 1993: 36 |
Pygathrix (Rhinopithecus) roxellana bieti:
| Groves 1970: 569 |
Rhinopithecus roxellanae bieti:
| Ellerman and Morrison-Scott 1951: 202 |
Rhinopithecus bieti
| : Milne-Edwards 1897: 156 |