
Gryllus transpecos Weissman & Gray, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4705.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:F534C43A-AB09-4CB3-9B08-FD5BDFD90298 |
|
persistent identifier |
https://treatment.plazi.org/id/182387A8-09B6-FF49-51F6-F93F05CAF829 |
|
treatment provided by |
Plazi |
|
scientific name |
Gryllus transpecos Weissman & Gray |
| status |
sp. nov. |
Gryllus transpecos Weissman & Gray , n. sp.
Texas Trans-Pecos Field Cricket
Figs 207–209 View FIGURE 207 View FIGURE 208 View FIGURE 209 , 216–221 View FIGURE 216 View FIGURE 217 View FIGURE 218 View FIGURE 219 View FIGURE 220 View FIGURE 221 , Table 1 View TABLE 1
‘G. #16’ and ‘G. #24’ of DBW notebooks.
Distribution. Only known from western Texas between the Rio Grande River and the Pecos River.
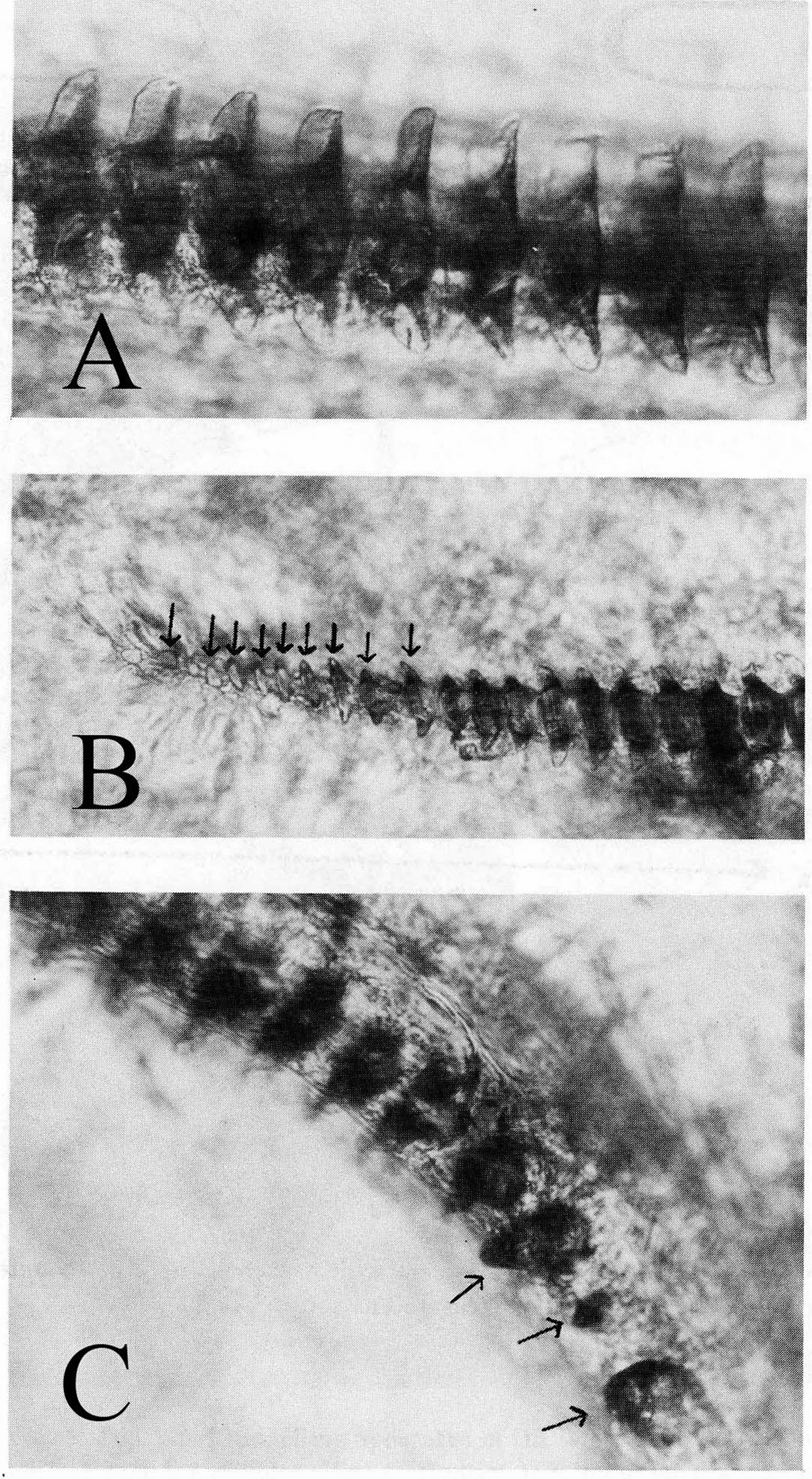
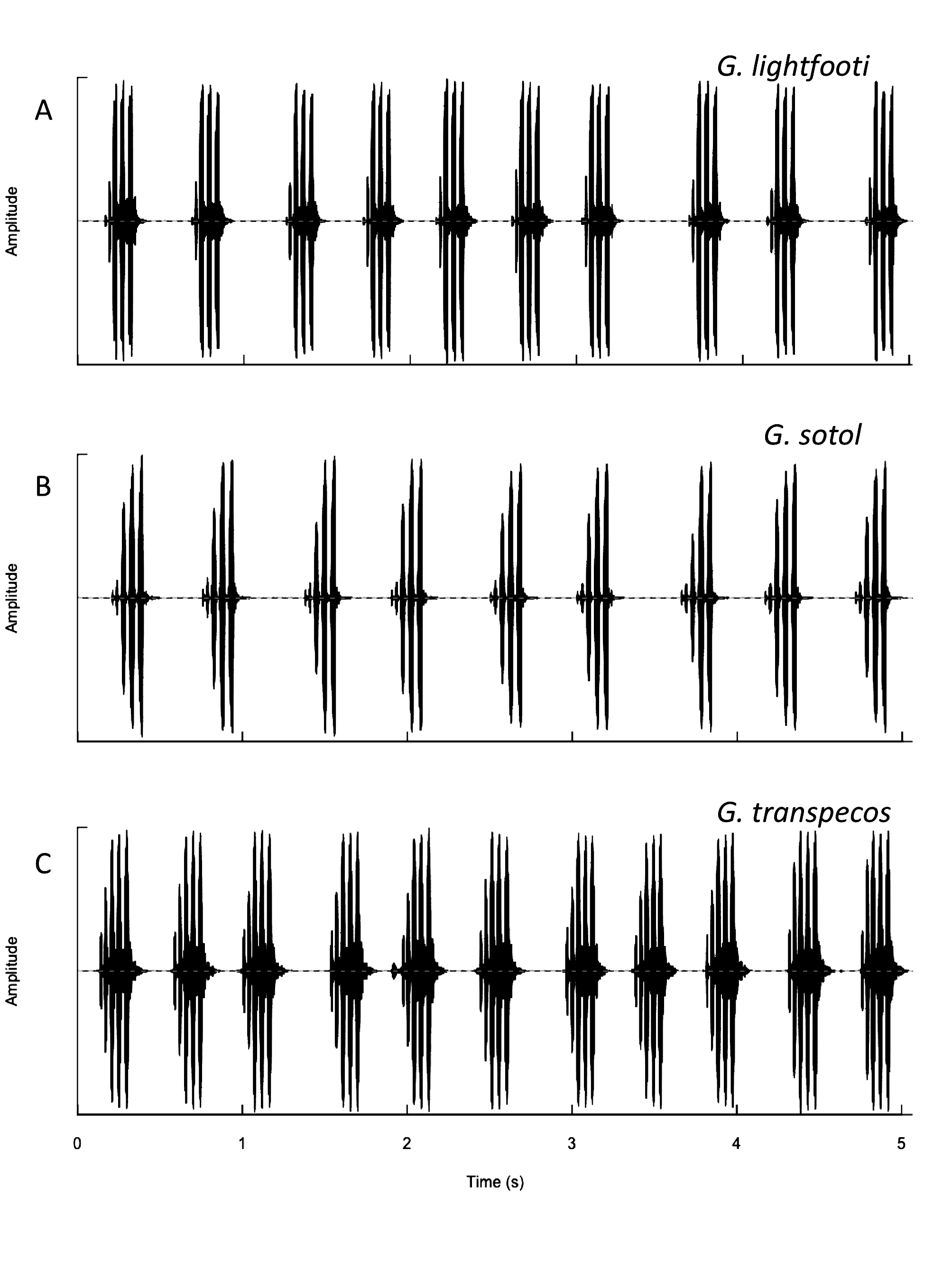
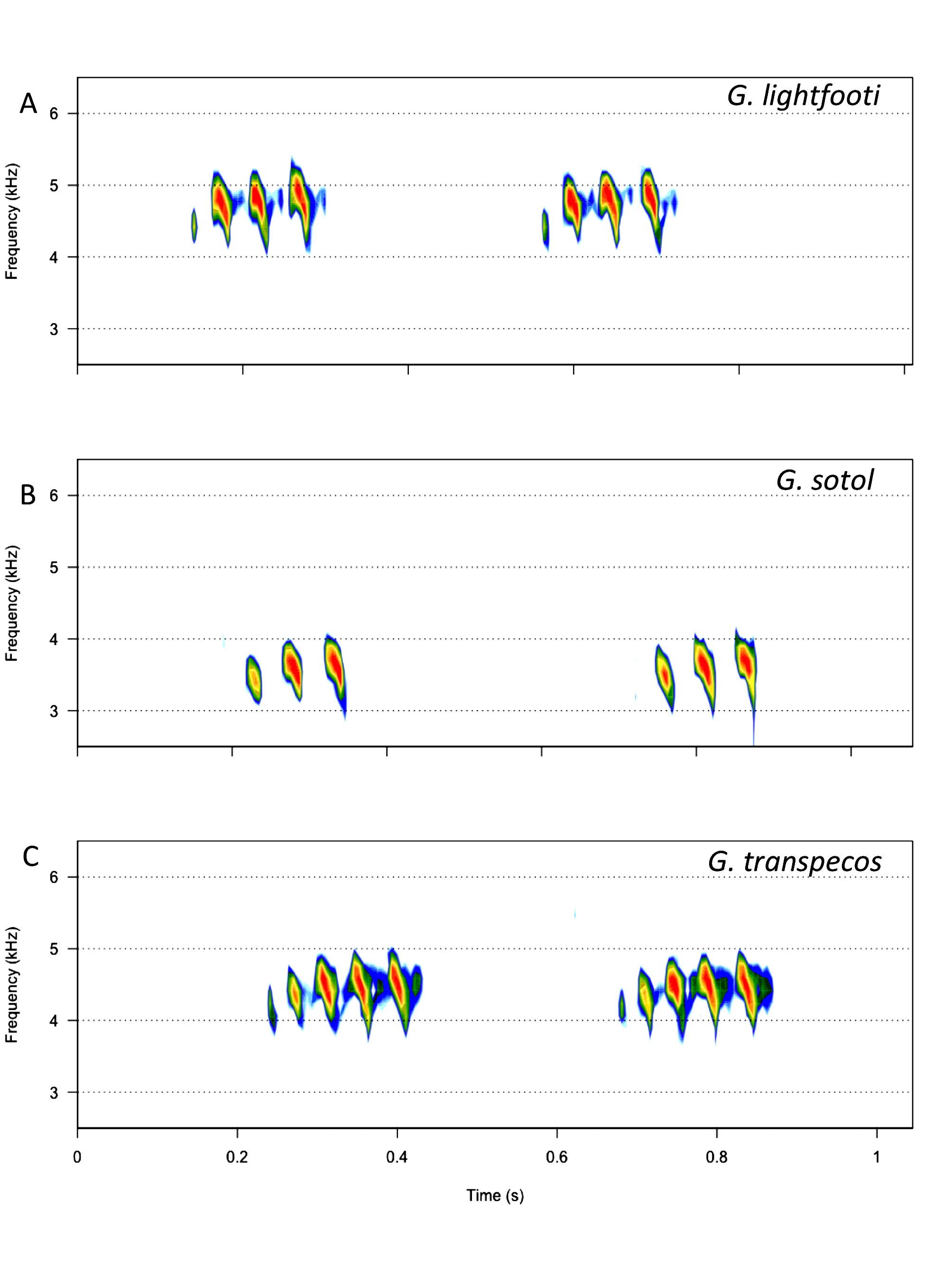
Recognition characters and song. Medium to large sized, wide head and pronotum, always short hind winged, female tegmina only cover ½ to ¾ of abdomen, cerci in situ typically extend beyond tip of ovipositor. Overall color black ( Fig. 216 View FIGURE 216 ) except for inside of hind femur which is reddish. Song ( Fig. 217 View FIGURE 217 , R09-93) a slow chirp of 4–5 (range 3–7) p/c, CR usually 80–150, PR 20–30 at 25°C. Closely related and similar to both G. lightfooti and G. sotol . Separated from G. lightfooti by, in G. transpecos , cerci in situ typically longer than ovipositor tip, female tegminal bars usually absent ( Fig. 216 View FIGURE 216 ), associated with rocks (vs. open grasslands), small but consistent ITS2 DNA differences ( Fig. 209 View FIGURE 209 ), and not known to climb into vegetation to sing. The two taxa could overlap in far western Texas, although no such locality presently known. From Van Horn Pass, where G. transpecos is common, to the town of Van Horn (S91-48), where G. lightfooti occurs, is some 4 miles distance. The allopatric, Organ Mts., NM, endemic and closely related, G. sotol ( Table 1 View TABLE 1 , p. 18) has more file teeth, higher tooth density, shorter cerci never as long as ovipositor tip (typically longer in G. transpecos ), slower PR, and different habitat: open, lower elevation, drier, rocky Chihuahuan Desert for G. transpecos versus higher, cooler, low mountain bajadas and woodlands, with sotol ( Dasylirion wheeleri ), for G. sotol . Additionally, G. sotol frequently climbs into sotol while G. transpecos is found on the ground, or sometimes on rocky cliff faces in road cut areas.
There are four other slow chirping, frequently rock-associated Gryllus species in the Trans-Pecos area that G. transpecos must be distinguished from. These four are not closely related to G. transpecos and can be separated by DNA (16S, ITS2, and multilocus data). Combinations of morphological, ecological and song characters are also diagnostic, as follows. From sometimes sympatric (at Van Horn Pass and Davis Mts.) G. longicercus : G. transpecos is distinguished by its larger head, broader pronotum, shorter cerci, lower tooth count and higher pulse rate. From almost sympatric (Davis Mts.) G. veletis : G. transpecos with longer cerci and different habitat (never in grassland). From sometimes sympatric (Davis Mts., S15-61) G. planeta , separated by 5 mostly non-overlapping physical characters ( Table 1 View TABLE 1 , p. 18): G. planeta has fewer number of teeth and shorter file length, cerci length, tegminal length and tegminal width, and faster chirp rate. From sometimes sympatric (at Big Bend, S85-55) G. chisosensis , they are separated by no overlap in number of teeth and PR and minor overlap in teeth/mm and cerci length. The two taxa are also separated by microhabitat in that G. chisosensis prefers rocky areas of many rock layers (e.g. talus slopes) while G. transpecos , while preferring rocky habitats like rocky road cuts, is not associated with extensive piles of rocks.
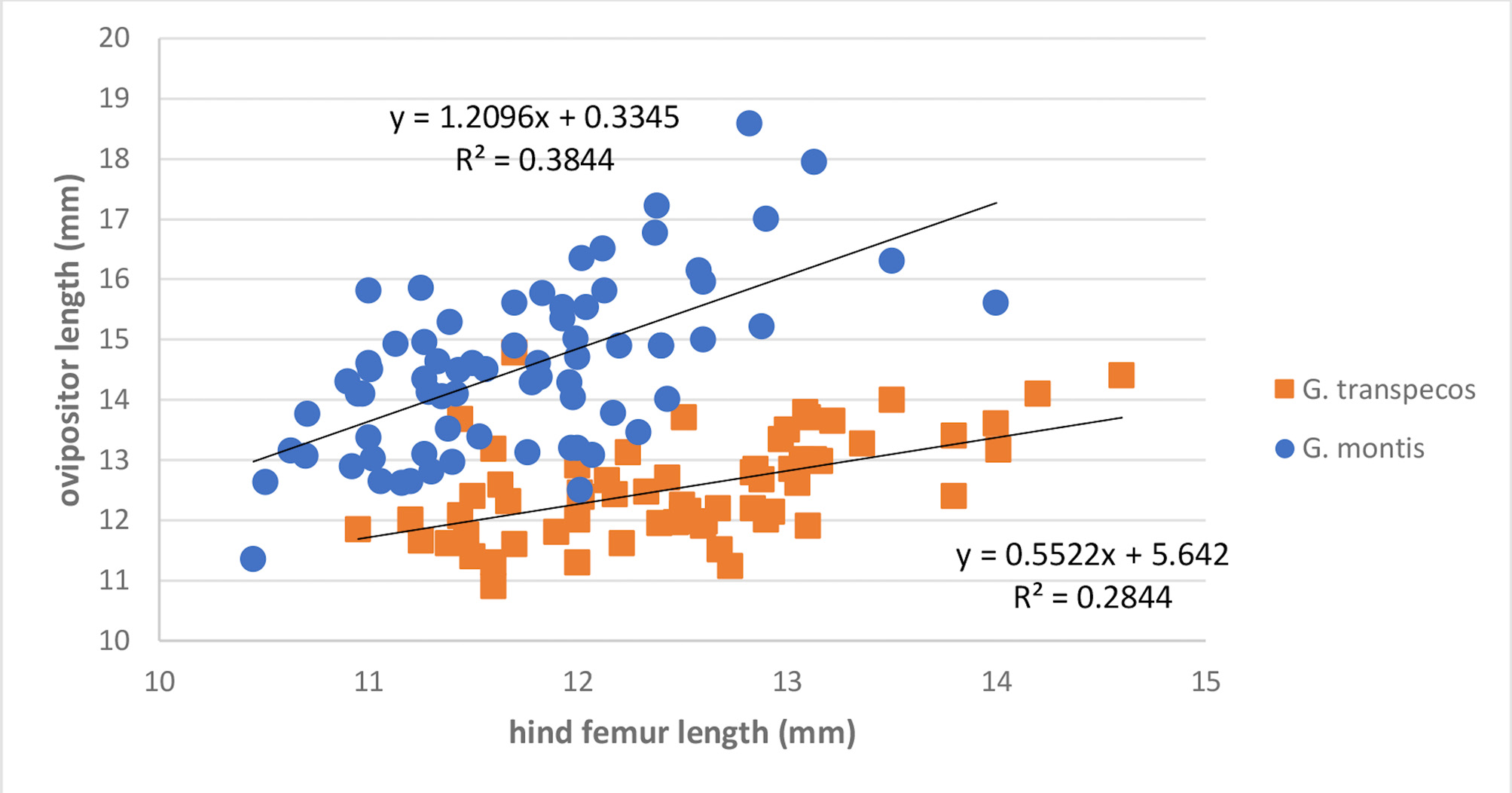
From other allopatric, widespread, more western and unrelated, slow chirping US Gryllus , G. transpecos is separated from G. montis which has shorter cerci, narrower head and pronotum, earlier appearance of adults, different habitats (latter almost always with an overhead tree canopy), and longer ovipositor ( Fig. 218 View FIGURE 218 ).
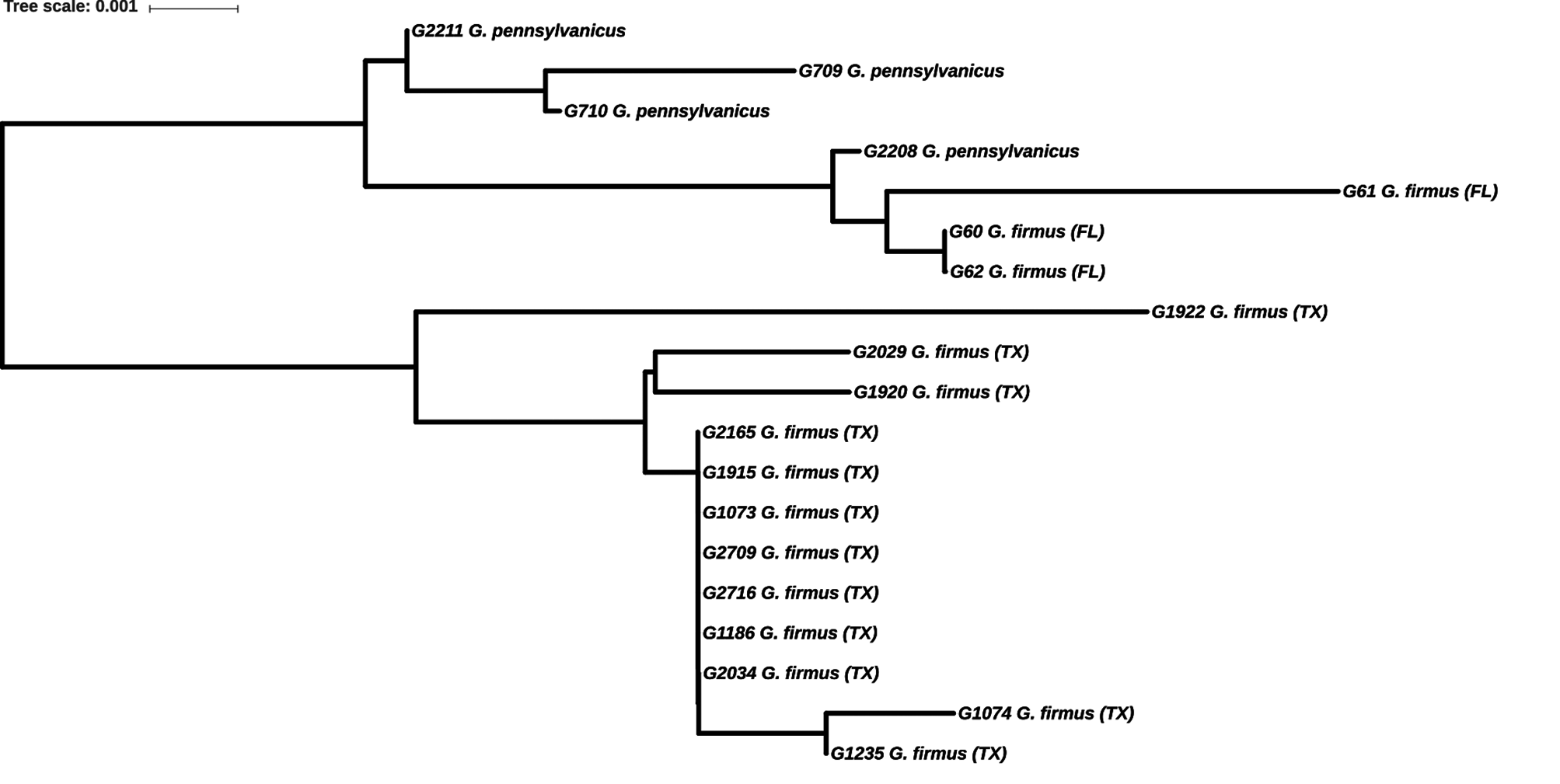
Separated by 800 km from more western G. saxatilis which has longer tegmina and longer ovipositor, different multilocus (Gray et al. 2019), 16S, and ITS2 ( Fig. 209 View FIGURE 209 ) DNA.
Holotype. Male ( Fig. 216 View FIGURE 216 ). USA, Texas, Brewster Co., Chisos Mts. , Big Bend National Park , Grande Village Road to Ranger Station near Lost Mine Trail parking lot, 1.5 m NW ranger station, 12-vi-2007. 5500’, 29° 16.471’ -103° 17.211’. D.B. Weissman & D.C. Lightfoot. S07-36, R07-95, G1175. 16S ribosomal RNA GenBank accession # MN 136663 View Materials . BL 19.49, HF 12.13, LC 12.36. Right tegmen removed: 142 teeth, file length 3.8, TL 11.9, TW 5.2. Deposited CAS, Entomology type #19273.
Paratypes. (Total: 105♂ 76♀). Texas, Brewster Co., Chisos Mts. , Big Bend National Park , 5500’, type locality, 8- vi-1985 (S85-55) 13♂ 6♀ ; 5-vi-1991 (S91-41) 11♂ 7♀ ; 12-vi-2007 (S07-36) 2♂ 1♀; Panther Junction , 3734’, 12-vi- 2007 (S07-38), 2-vii-2015 (S15-68) 3♂ 3♀ ; 18.7 m SE Panther Junction , 9-vi-1985 (S85-57) 1♂ ; 5-vi-1991 (S91-42) 1♂ ; Maverick Rd. , 3756’, 2-vii-2015 (S15-69) 2♂ ; Rio Grande Village , 2100’, 9-vi-1985 (S85-56) 4♂ 1♀ ; 5-vi-1991 (S91-43) 2♂ ; Grande Village Road 4.5 m NW ranger station, 4340’, 12-vi-2007 (S07-37) 1♂ 3♀ ; 28-v-2016 (S16- 12) 1♂ . Hwy 118 24 m S Alpine, 4434’, 2-vii-2015 (S15-72) 7♂ 10♀ . Hwy 170 8 m W Terlingua, 3000’, 27-v-2016 (S16-10) 1♂ . Hwy 385 3.3 m S Marathon, 4050’, 28-v-2016 (S16-14) 2♂ 1♀ . Culberson Co., Van Horn Pass area , 4880’, 31° 03’ 17.0” -104° 57’ 04.4”, 12-vi-1985 (S85-65) 6♂ 4♀ ; 6-vi-1991 (S91-47) 11♂ 11♀ ; 29-vi-2009 (S09-64) 10♂ 3♀. Jeff Davis Co., Davis Mts., Observatory , 6747’, 1-vii-2015 (S15-61) 4♂ 1♀ ; 2.9 road m below Observatory , 6010’, 1-vii-2015 (S15-64) 1♂ 2♀ ; 3.2 road m below Observatory , 5935’, 1-vii-2015 (S15-65) 5♂ 5♀ . Presidio Co., Chianti Mts., Hwy 67 Shafter , 3880’, 27-v-2016 (S16-3) 4♂ ; 0.5 m N Shafter , 3856’, 27-v-2016 (S16-2) 7♂ 15♀ ; 2.3 m S Shafter , 26-vi-1986, 3900’ (S86-45) 3♂ 2♀ . Hwy 170 E. Presidio , 2680’, 27-v-2016 (S16-7) 1♂ 1♀ . Hwy 170 39 m W Terlingua, 2660’ 27-v-2016 (S16-8) 1♂ ; 30 m W Terlingua , 2600’, 27-v-2016 (S16-9) 1♂ .
Derivation of name. Trans-Pecos was originally defined, in 1887, by Texas geologist Robert T. Hill, as that portion of Texas west of the Pecos River, which perfectly describes the distribution of this species.
Geographical range. ( Fig. 219 View FIGURE 219 ). Apparently absent from ecologically similar areas of the Guadalupe Mts., Culberson Co., Texas, where we found (S01-63, 64, 65) slow chirpers G. longicercus and G. lightfooti at lower, dryer elevations, and G. veletis at higher (1920m), more mesic Dog Canyon.
Habitat. Rocky road cuts (S16-2, 16-4); Piñon-juniper-oak woodland (S07-36); Chihuahuan Desert (S07-37) with Acacia , sotol, and Yucca ; and natural, rocky slopes in and around towns (S16-3). Males sing from crevices, holes, under small plants, and under and around rocks. Not known from within any town, where G. lightfooti can frequently occur. G. transpecos appears to be the ecological equivalent of more western G. saxatilis .
Life cycle and seasonal occurrence. No egg diapause: Big Bend (S85-55 and S85-56), Van Horn (S85-65). Probably one generation/year. No nymphs seen at any Brewster Co. locality but most of population south of Shafter (Presidio Co., S86-45) penultimate or last instar when collected on 26-vi-1986.
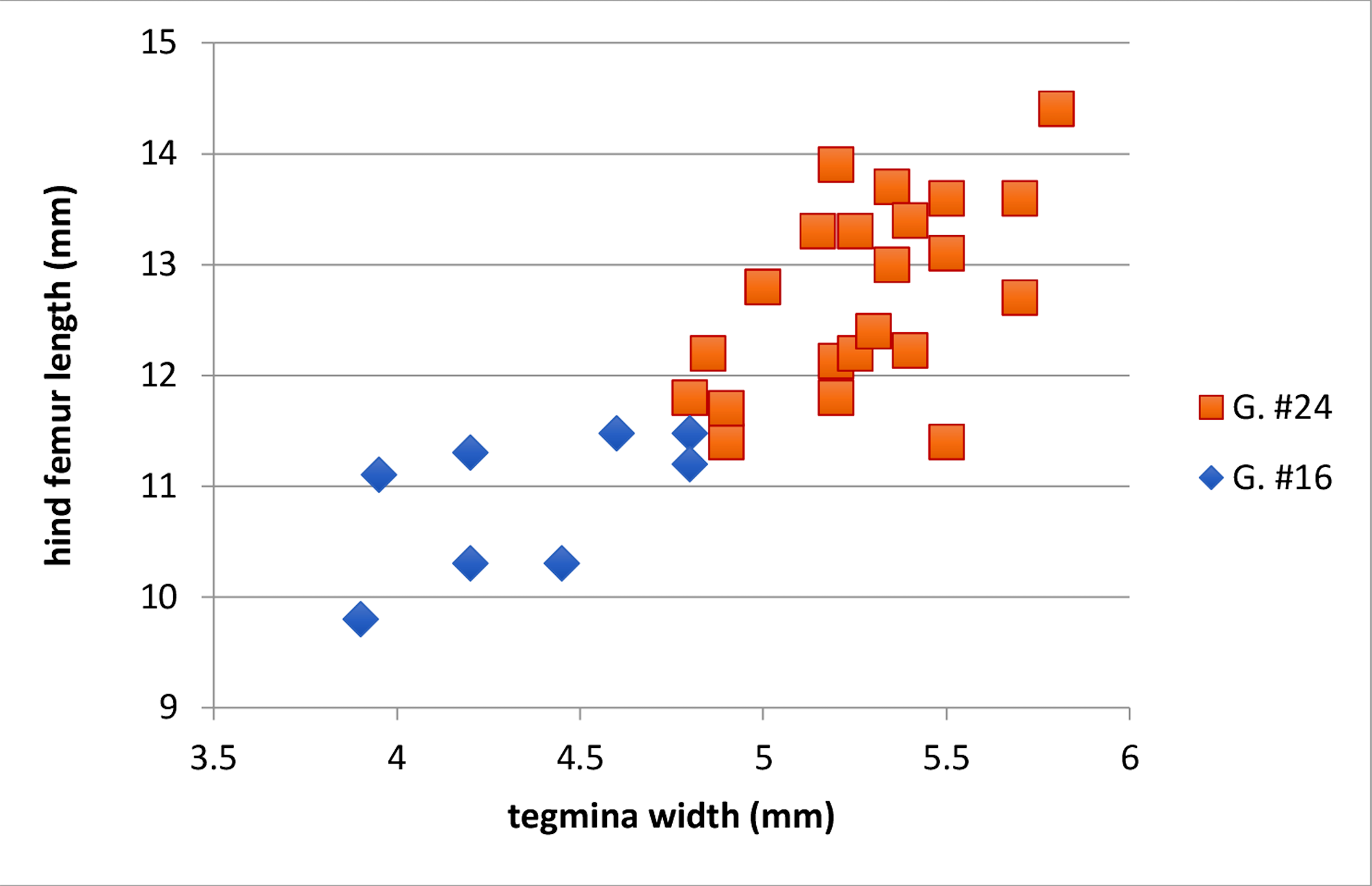
Variation. Specimens from Big Bend National Park (our ‘G. #24’) are larger than those from Van Horn Pass (our ‘G. #16’) (see Table 1 View TABLE 1 , p. 18) and this caused us some initial confusion. Individuals from these two localities differed in temperature corrected pulse rate, with #16 slower than #24 (21 ± 0.4 p/s versus 24 ± 0.4 p/s; F 2,22 = 12.02, P <0.001)); males differed in teeth/mm vs. hind femur length ( Fig. 220 View FIGURE 220 ; F 2 View FIGURE 2 , 30 View FIGURE 30 = 5.5, P <0.01) and tegmina width vs. hind femur length ( Fig. 221 View FIGURE 221 ; F 2 View FIGURE 2 , 30 View FIGURE 30 = 7.4, P <0.01). There was no difference in chirp rates, pulses/chirp, or dominant frequency. However, with the addition of individuals from areas other than these two localities, these significant differences disappeared, and we currently attribute this variation mostly to differences in body size, which is consistent with the linear relationships seen in both regressions—i.e. larger crickets have larger body proportions.
DNA. Multilocus G3062 Big Bend National Park (S15-68) and G3083 Davis Mts. (S15-61) sister species of G. sotol and G. lightfooti (Gray et al. 2019) . We note that our confusion re. G. #16 and G. #24 (see above) was compounded by the fact that early 16S sequencing of G. transpecos yielded 3 separate clades comprised of individuals from 5.3 km south Marathon (S16-14), individuals from 38.6 km south of Alpine (S15-72), and individuals from Van Horn Pass (S09-64). Subsequent ITS2 sequencing showed these same individuals, from all three sites, to cluster together ( Fig. 209 View FIGURE 209 , p, 207 View FIGURE 207 ), hence our combining them together under G. transpecos .
Discussion. Can be a relatively common field cricket where found. Singing males easy to approach but stop singing once turning over rocks near them, as opposed to many males of G. longicercus that continue to sing while “their” rocks being turned over. Can sing all night and into early morning in Big Bend (S85-55, at 08:30).
Tinkham (1948) discussed the Orthopteran and faunistic zones of the Big Bend Region.



| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.