
Pterosphenus cf. schweinfurthi ( Andrews, 1901 )
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2021v43a5 |
|
publication LSID |
urn:lsid:zoobank.org:pub:697FC553-E37B-4EF9-97A4-950E4DEE246C |
|
DOI |
https://doi.org/10.5281/zenodo.4606645 |
|
persistent identifier |
https://treatment.plazi.org/id/03923C45-FF91-FF99-30DB-FACCFBEF1185 |
|
treatment provided by |
Felipe |
|
scientific name |
Pterosphenus cf. schweinfurthi ( Andrews, 1901 ) |
| status |
|
Pterosphenus cf. schweinfurthi ( Andrews, 1901)
EXAMINED MATERIAL. — Four vertebrae recovered from the Garouaze Locality in Gueran Depression. Two of the specimens (FSAC Bouj -300 and 317) clearly belong to the Palaeophiidae . The two other vertebrae (FSAC Bouj -316 and 318) are very incomplete, but they also likely belong to palaeophiids.
DESCRIPTION
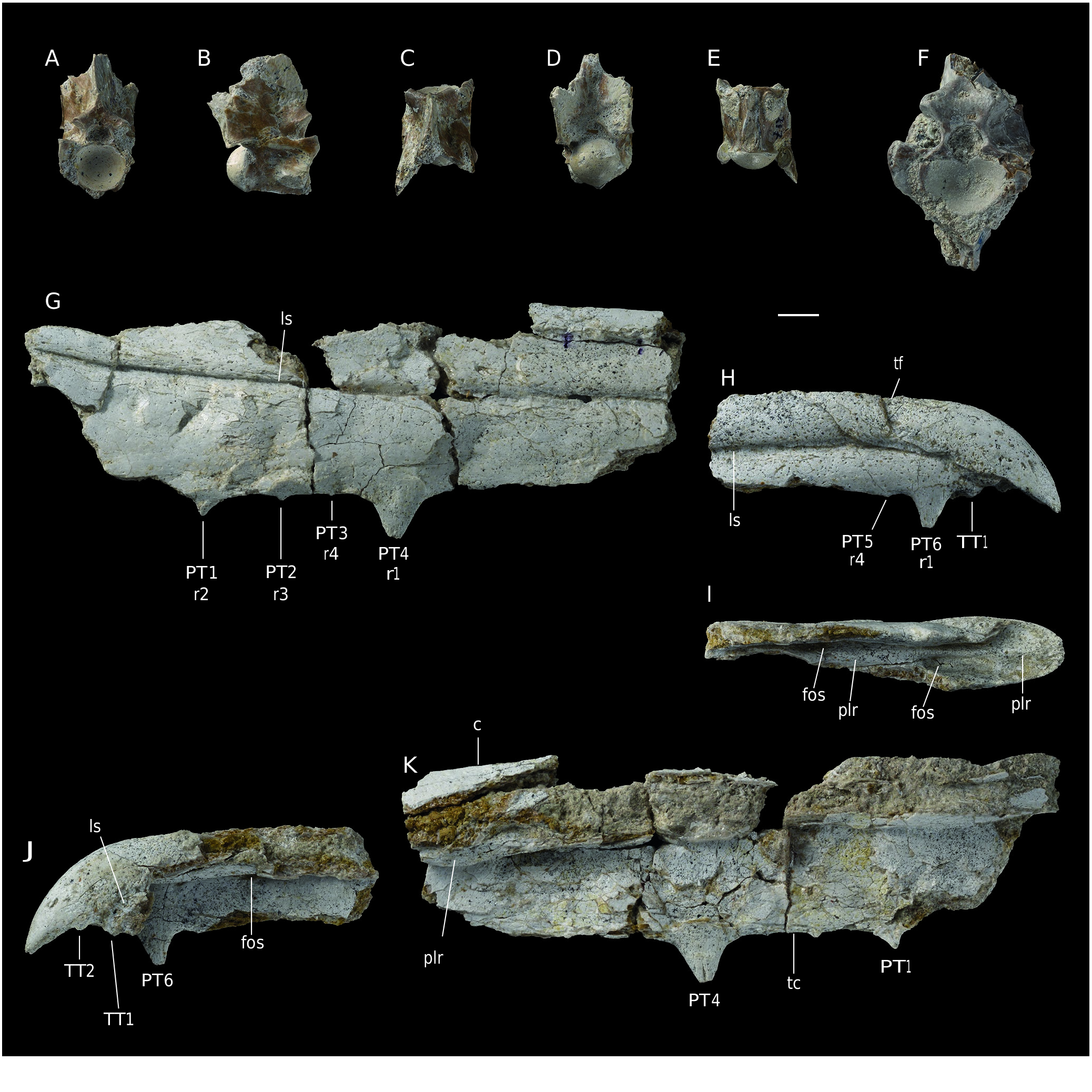
The description is mainly based on FSAC Bouj-317 ( Fig. 6 View FIG A-E) but more information is drawn from FSAC Bouj-300 ( Fig. 6F View FIG ). These vertebrae belong to a large snake. In FSAC Bouj-317, measurements are as follows: centrum length, from the cotyle rim to the tip of condyle = 21.7 mm; horizontal diameter of cotyle = 13.1 mm; zygosphene width = 13.7 mm. FSAC Bouj-300 is larger, but its centrum length cannot be measured; the horizontal diameter of its condyle is approximately 19.9 mm and the width of its zygosphene is 21.6 mm. Both vertebrae are tall, short and compressed laterally. FSAC Bouj-317 preserves incomplete pterapophyses above the postzygapophyses.
In anterior view, FSAC Bouj-317 is clearly compressed laterally, and is very narrow. The cotyle is broad and approximately circular but somewhat truncated dorsally. The size of the preserved prezygapophysis, on the right side, appears much reduced compared to the cotyle. The neural canal is small. The articular facet of the prezygapophysis lies approximately at the level of the floor of the neural canal. The width of the zygosphene is nearly similar to that of the cotyle. The zygosphene is relatively thick and arches dorsally. The top of the zygosphene forms the base of the anterior border of the neural spine. FSAC Bouj -300 differs from FSAC Bouj- 317 in being slightly less compressed laterally and in having a markedly thicker zygosphene.
In dorsal view, FSAC Bouj-317 appears narrow and comparatively elongate. The interzygapophyseal constriction is so shallow that it is almost not expressed. The axis of the small prezygapophyseal facet is directed anterolaterally. The anterior border of the zygosphene forms an obtuse notch. The neural spine extends through the whole preserved length of the neural arch; anteriorly, it reaches the anterior border of the zygosphene. FSAC Bouj-300 was likely less narrow and less elongate.
In lateral aspect, the neural spine is long anteroposteriorly; unfortunately, its dorsal part is broken away and its height cannot be estimated. Its anterior border comprises a vertical portion that rises from the zygosphene and a longer, posteriorly inclined dorsal portion. The vertebra does not preserve the tips of both prezygapophyses, so that the length of the latter remains unknown. The anterior edge of the pterapophyses is inclined posterodorsally at an angle of approximately 45°. The interzygapophyseal ridge is very prominent but blunt. The prezygapophyseal buttress forms an anterolateral sharp ridge. Unfortunately, the two paradiapophyses are completely eroded and hypapophyses are not preserved. The axis of the condyle is horizontal. FSAC Bouj-300 only provides one additional information: it bears the basis of a vertical hypapophysis. It is not possible to determine whether or not an anterior hypapophysis was present.
The ventral face of the centrum of FSAC Bouj-317 is narrow and elongate, not limited by subcentral ridges. The sagittal area is damaged. The bases of the paradiapophyses are markedly separated from each other. The ventral face of FSAC Bouj-300 is poorly preserved. However, its sagittal area forms a carina. The hypapophysis originates from the posterior portion of this carina. Apparently, there is no room for an anterior hypapophysis.
The posterior aspect of FSAC Bouj-317 is striking. Above the neural canal, the neural arch is extremely thick and bounded laterally by vertical borders. The dorsolateral parts of the neural arch form the bases of the broken pterapophyses. The posterior face of FSAC Bouj-300 does not display observable characters.
FSAC Bouj-316 and 318 are two centra whose morphology is consistent with those of FSAC Bouj-300 and 317. Although very incomplete, both specimens bear an entirely preserved hypapophysis. In both vertebrae, the short, laterally compressed hypapophysis shows a vertical posterior border that contacts the condyle and a weakly (in FSAC Bouj-318) or strongly (in FSAC Bouj-316) inclined anterior border. It is not possible to determine whether an anterior hypapophysis was present in these two vertebrae.
COMPARISON AND REMARKS
The lateral compression, reduced prezygapophyses, prezygapophyseal buttresses forming an anterolateral edge, presence of pterapophyses and horizontal axis of condyle constitute a combination of characters that occur only in Palaeophiidae . In addition, the shortness and height of the vertebrae, as well as low position of the zygapophyseal plane, make it possible to discard the Archaeophiinae and to refer the specimens to the Palaeophiinae ( Rage et al. 2003) .
Two genera, Palaeophis Owen, 1841 and Pterosphenus , are assigned to the Palaeophiinae of Palaeophiidae , a family including species of various sizes, slightly adapted to strongly specialized for aquatic life, and widely distributed from America to Asia. The species from Gueran is large and marine, and it was first identified in Fayum ( Andrews 1901). The vertebrae of the species referred to these two genera form a morphological cline ( Rage 1983a). Compared with the generalized snake vertebrae, Palaeophis species have vertebrae displaying the more conservative morphology. In Pterosphenus , vertebrae are more laterally compressed, the prezygapophyses are more reduced, and the pterapophyses are taller than in Palaeophis . However, in the morphocline, there is a blurred transition between species that may be referred to either Palaeophis or Pterosphenus ; the distinction between the two genera is phenotypic and artificial ( Rage 1983a).
Fortunately, the morphology of the vertebrae from Gueran is consistent with that of the more derived species. Consequently, the specimens are assigned to Pterosphenus . This assignment based on the degree of lateral compression and height of the vertebrae is confirmed by the continuity between the top of the zygosphene and the anterior border of the neural spine; this feature is known only in Pterosphenus ( Rage 1983b) . This character unquestionably occurs in FSAC Bouj-300 and FSAC Bouj-317 (it cannot be checked in FSAC Bouj-316 and FSAC Bouj-318). The zygosphene of FSAC Bouj-300 is markedly thicker and narrower than that of FSAC Bouj-317, which corresponds to intracolumnar variation. FSAC Bouj-300 is a vertebra from the mid-trunk portion whereas FSAC Bouj-317 comes from the posterior trunk region. The smaller size of FSAC Bouj-317 likely also reflects intracolumnar variation; the anterior and posterior trunk regions of palaeophiids appear to have been slenderer than the mid-trunk portion. FSAC Bouj-300 and 317 likely belong to the same species (but no conclusion can be made about 316 and 318).
Five recognized species belong to Pterosphenus ( Rage et al. 2003; McCartney & Seiffert 2016): P. schucherti , the type species, middle (Bartonian and?Lutetian) and late (Priabonian) Eocene of the United States; P. schweinfurthi , late Eocene (Bartonian) of Libya and late Eocene (Priabonian) of Egypt: note that McCartney & Seiffert (2016) assigned an Eocene/Oligocene age to the fossils from Libya; however, we follow Abouessa et al. (2012) who referred the Libyan locality to the late Bartonian. P. sheppardi , late Eocene of Ecuador; P. kutchensis and P. biswasi , both from the lower Eocene (Ypresian) of India. The fact that paradiapophyses are clearly separated from each other and that the anterior border of the neural spine reaches the anterior border of the zygosphene allows to exclude P. kutchensis . The morphology of FSAC Bouj-300 and 317 is close to that of P. biswasi . However, the latter species differs from FSAC Bouj-300 and 317 in having a less concave anterior border of the zygosphene. Another possible difference is that, on the centrum, the base of each paradiapophysis is less extended dorsoventrally in FSAC Bouj-317 (not observable in FSAC Bouj-300) than in P. biswasi . Comparison with P. sheppardi is difficult because this species is only represented by five articulated vertebrae. If the vertebrae were articulated, several significant characters would have been concealed. However, the pterapophyses of P. sheppardi are smaller and shorter than those of FSAC Bouj-317 (no possible comparison with FSAC Bouj-300). If this difference is not an intracolumnar variation, then it is significant at the species level variations. Distinction between P. schucherti and P. schweinfurthi is not clear. Specific differences that were put forward are perhaps only intracolumnar variation ( Rage et al. 2003; Parmley & Devore 2005; McCartney & Seiffert 2016). Pterosphenus schweinfurthi may be a junior synonym of P. schucherti but this cannot be demonstrated. FSAC Bouj-300 and 317 do not show significant differences with the known vertebrae of both P. schucherti and P. schweinfurthi ( Lucas 1899; Janensch 1906; Westgate & Ward, 1981; McCartney & Seiffert 2016). However, in view of the limited material and its incomplete nature and taking into account the uncertainty that remains about the possible synonymization of P. schweinfurthi with P. schucherti , we assign the material from Gueran to Pterosphenus cf. schweinfurthi .
It is worth noting that the geographically close locality of Ad-Dakhla yielded some palaeophiid vertebrae. Bedbone 1 from which the palaeophiid fossils in Ad-Dakhla area came from is slightly younger than the fossiliferous level of Gueran. Zouhri et al. (2014) assigned a Priabonian age to Bedbone 1. In the vertebrae from Ad-Dakhla, the junction between the anterior borders of the zygosphene and neural spine clearly belongs to the Pterosphenus type. In addition, one (unnumbered) incomplete vertebra displays proportions that are similar to those of the known vertebrae of the P. schweinfurthi - P. schucherti assemblage and may be referred to P. cf. schweinfurthi . However, one vertebra from Ad-Dakhla (Dak-349) is less compressed laterally and its proportions resemble those of species belonging to the transition between Palaeophis and Pterosphenus . It is not possible to state whether Dak-349 belongs to a species distinct from the P. schweinfurthi - P. schucherti assemblage or if it represents an intracolumnar variation of the later assemblage that was hitherto unknown. Whatever the case may be, Pterosphenus cf. schweinfurthi is present in Gueran (Bartonian) as in Ad-Dakhla.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Elasmobranchii |
|
SuperOrder |
Batoidea |
|
Order |
|
|
Family |
|
|
Genus |