
Scyliorhinus hachijoensis, Ito & Fujii & Nohara & Tanaka, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5092.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:176F682C-17CF-4E86-9414-DCA5DE254D91 |
|
DOI |
https://doi.org/10.5281/zenodo.5889092 |
|
persistent identifier |
https://treatment.plazi.org/id/0382EA3B-FFCA-FFEB-0FC7-8909FCF0CF90 |
|
treatment provided by |
Plazi |
|
scientific name |
Scyliorhinus hachijoensis |
| status |
sp. nov. |
Scyliorhinus hachijoensis sp. nov.
( Table 1 View TABLE 1 and Figs. 1–9 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
New English name: Cinder cloudy catshark; new Japanese name: Fukami-torazame.
Scyliorhinus sp. : Hagiwara, 1993: 1, 9, Tables 1 View TABLE 1 –3, Fig. 3 View FIG (keeping and reproduction)
Holotype.
NSMT-P135960 , adult male, 370 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 200–300 m depth, 24 October 2018).
Paratypes.
MSM-19-292, female, 331 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300–400 m depth, 12 July 2018); MSM-19-293, female, 354 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300– 400 m depth, 12 July 2018); MSM-19-294, female, 326 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300–400 m depth, 12 July 2018); MSM-19-295, female, 354 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300–400 m depth, 12 July 2018); MSM-19-296, female, 368 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300–400 m depth, 12 July 2018); MSM-19-297, female, 382 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300–400 m depth, 12 July 2018); MSM-19-298, female, 366 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 200–300 m depth, 18 August 2018); MSM-19-299, female, 350 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 200–300 m depth, 18 August 2018); MSM- 19-300, female, 356 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 200–300 m depth, 18 August 2018); MSM-19-301, female, 368 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 200–300 m depth, 18 August 2018); NSMT-P 135961, female, 322 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 400–500 m depth, 12 July 2018); NSMT-P 135962, female, 348 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 500 m depth, 11 July 2018); NSMT-P 135963, male, 384 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); NSMT-P 135964, female, 364 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); NSMT-P 135965, male, 419 mm TL (off the east coast of Torishima Island, Tokyo, Japan, 600–650 m depth, 27 June 2018); NSMT-P 136476, female, 386 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300 m depth, 15 November 2018); NSMT-P 136477, female, 342 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); NSMT-P 136478, male, 346 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45542, female, 370 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300 m depth, 15 November 2018); SPMN-PI 45543, female, 364 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300 m depth, 15 November 2018); SPMN-PI 45544, female, 345 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300 m depth, 15 November 2018); SPMN-PI 45545, female, 372 mm TL (off the east coast of Hachijojima Island, Tokyo, Japan, 300 m depth, 15 November 2018); SPMN-PI 45546, female, 301 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45547, female, 355 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45548, female, 305 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45549, male, 294 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45550, male, 335 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018); SPMN-PI 45551, female, 356 mm TL (off the east coast of Mikurajima Island, Tokyo, Japan, 100–200 m depth, 24 October 2018).
Other materials.
HUMZ113575, female, 353 mm TL (off Torishima Island, Tokyo, Japan); HUMZ113576, male, 372 mm TL (off Torishima Island, Tokyo, Japan).
Diagnosis.
A species of Scyliorhinus distinguished by its anterior nasal flaps not reaching the upper lip (vs. flaps reaching upper lip, and sometimes covering it, in S. canicula , S. cervigoni , S. comoroensis , S. duhamelii , S. garmani and S. stellaris ); nasoral grooves absent and posterior nasal flaps situated posterior to excurrent apertures (vs. nasoral grooves prexents and posterior nasal flaps laterally situated in S. canicula and S. duhamelii ); mouth length less than half of mouth width (vs. mouth length more than or equal half of mouth width except in S. torazame and S. ugoi ); anal fin height more than caudal peduncle height (vs. less than caudal peduncle height in S.boa , S. duhamelii , S. torazame and S. torrei ), and greater than or equal to half of mouth width (vs. less than half of mouth width in S. boa , S. capensis , S. duhamelii , S. haeckelii , S. hesperius , S.meadi , S. torazame , S. torrei and S. ugoi ); saddles darker than the background color (vs. inconspicuous or absent in S. boa , S. cabofriensis , S. cervigoni , S. duhamelii , S. garmani and S. torrei , and dark lines in S. retifer ); body grayish brown to dark brown with well-defined light spots and small dark spots (vs. spots absent in S. retifer , yellow to golden spots in S. capensis , light spots absent in S. cervigoni , S. garmani , S. meadi and S. retifer , and dark spots absent in S. capensis , S. comoroensis , S. hesperius , S. meadi , S. torazame and S. torrei ); light spots spiracle-sized or larger (vs. predominantly smaller than spiracles in S. boa , S. cabofriensis , S. canicula , S. duhamelii , S. stellaris and S. ugoi ); dark spots smaller than spiracles (vs. predominantly larger than spiracles in S. cervigoni , S. duhamelii , S. garmani , S. haeckelii and S. stellaris ); number of monospondylous vertebrae 34–36 (vs. counts higher except in S. duhamelii , S. torazame and S. torrei ); clasper with hooks (vs. absent in all other species except S. torazame ); accessory terminal cartilage present (vs. absent in S. cabofriensis , S. cervigoni , S. comoroensis , S. duhamelii , S. haeckelii , S. stellaris , S. torrei and S. ugoi ); egg case surface with irregularities (vs. smooth in all other species).
Description.
Morphometric measurements are given in Table 1 View TABLE 1 .
Body slender and tapering to caudal fin ( Fig. 1 View FIG ), precaudal length 77.3 %TL (72.0–76.4 %TL in palatypes). Prepectoral length 0.4 times prepelvic length. Pectoral–pelvic space 1.5 times pelvic–anal space (1.7–2.5 times in female, 1.4–1.9 times in male). Interdorsal space 1.6 times dorsal–caudal space. Trunk shorter than tail; snout–vent length 44.1 %TL (42.0–44.6 %TL). First dorsal origin above insertion of pelvic fin, second dorsal origin forward to anal insertion. No interdorsal, postdorsal, or postanal ridges; lateral crest on caudal peduncle absent.
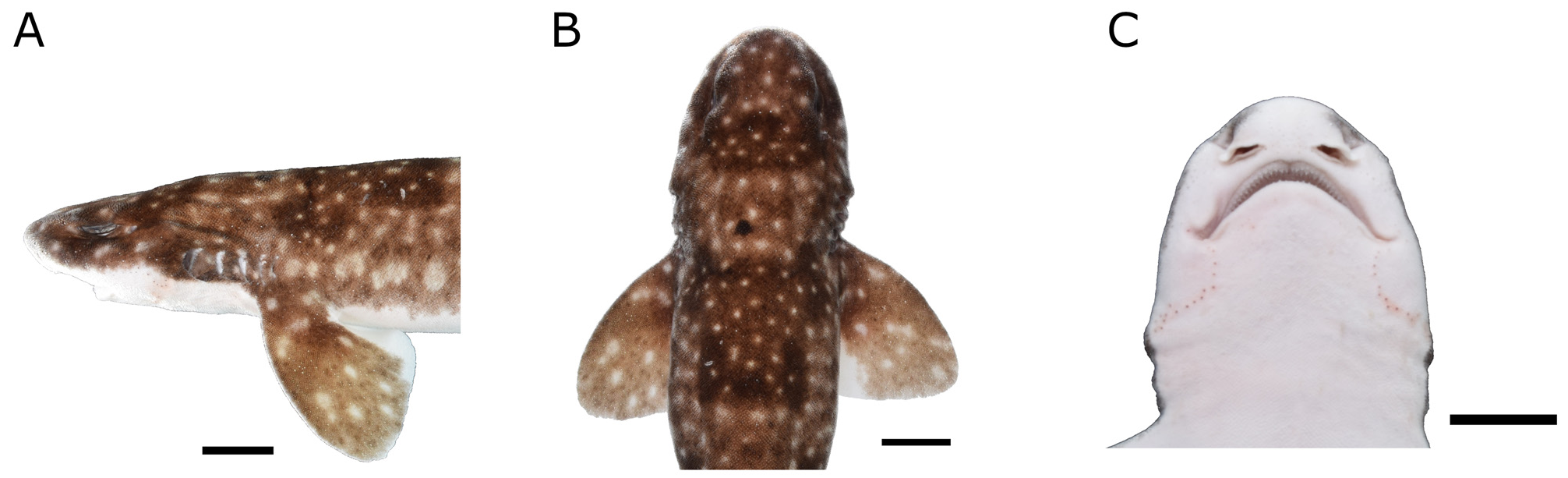
Head ( Fig. 2 View FIG ) moderately broad and depressed. Head length (HL) 18.9 %TL (17.2–19.6 %TL) and 1.4 times head width. Snout short, prenasal length 2.3 %TL (2.1–2.6 %TL) and 0.6 times preoral length. Preoral length 3.8 %TL (3.2–4.4 %TL), 0.4 times mouth width and 0.9 times preorbital length. Preorbital length 4.1 %TL (4.1–5.1 %TL), 0.5 times interorbital space.
Eyes ( Fig. 2A View FIG ) large and slitlike, eye length 4.1 %TL (3.3–4.4 %TL), 0.2 times head length, with lower edges medial to horizontal head rim in dorsal view; subocular ridge strong. Nictitating lower eyelid of rudimentary type, with shallow subocular pocket and secondary lower eyelid free from upper eyelid. Eye length 5.3 times and interorbital distance 9.6 times spiracle length.
Spiracles ( Fig. 2A View FIG ) close behind but clearly separated from eyes, positioned dorsolaterally on head and somewhat lower than level of eye notch.
First and second gill openings about equal in height, with last three tapering down to fifth ( Fig. 2A, B View FIG ). First gill opening height 1.6 times that of fifth. All gill openings slightly concave and not elevated on dorsolateral surface of head. Gill filaments not visible externally.
Nostrils ( Fig. 2C View FIG ) with broad incurrent apertures, without nasoral grooves or nasal barbels and with small and oval excurrent apertures. Anterior nasal flaps large, triangular, and covering posterior nasal flap and excurrent aperture, extending just anterior to mouth, close to but not touching upper lip. Mesonarial ridge distinct but not exceeding posterior border of anterior nasal flap. Posterior nasal flaps small and rectangular, situated on posterior border of excurrent aperture. Internarial distance 0.3 times interorbital distance.
Mouth ( Fig. 2A, C View FIG ) arched, moderately wide, and short. Mouth length 2.9 %TL (2.5–4.4 %TL), 0.3 times mouth width and 0.2 times head length; mouth width 0.7 times head width. Upper labial furrows absent. Lower labial furrows short and narrow, length 0.1 times mouth width.
Tooth counts 39/36 (36–49/ 32–43 in paratypes); teeth morphologically similar in both jaws. Monognathic heterodonty gradual and well developed; anterolateral teeth abruptly larger than symphysial ones and smaller distally; posterior teeth with smaller principal cusps than anterolateral teeth ( Fig. 3 View FIG ). Sexual heterodonty weak. Symphysial teeth with principal cusp and one weak cusplet on each side. Anterolateral teeth with more developed principal cusp than symphysial teeth and two to four cusplets; most outside cusplets small and undeveloped when with three or four cusplets. Posterior teeth have two or three cusplets and tilted to the outside; principal cusp weakly developed.
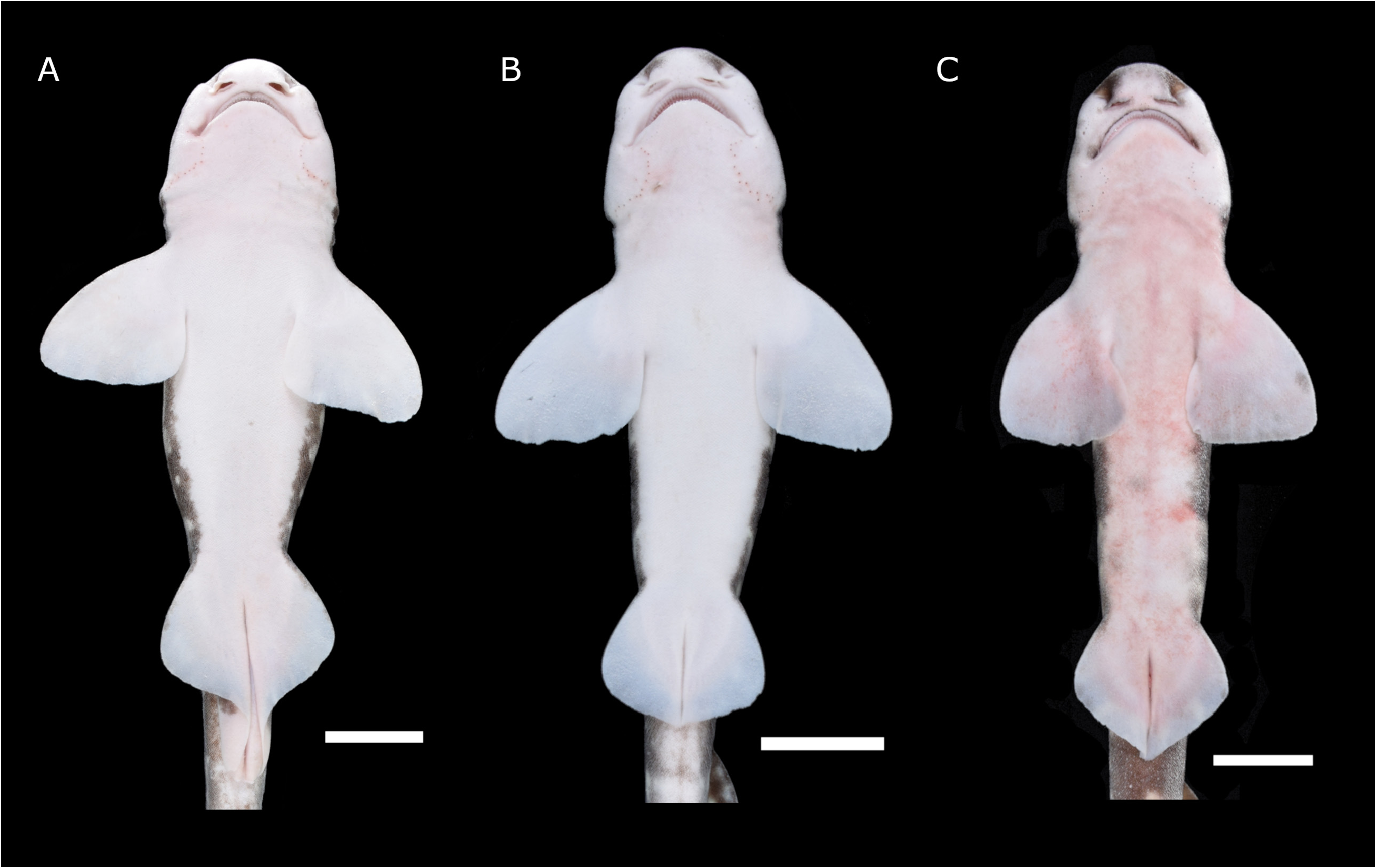
Pectoral fins ( Figs. 1 View FIG , 4A, B View FIG ) large and rounded-triangular, not falcate, with broadly convex anterior margins and almost straight posterior and inner margins. Pectoral fin inner corner obtuse angle (96º in holotype, 90º–102º in paratypes). Pectoral origin under third gill opening, with base length 0.8 times mouth width. Pectoral anterior margin 2.4 times base length and 1.5 times posterior margin.
Pelvic fins ( Figs. 1 View FIG , 4A, B View FIG ) broadly triangular, with rounded outer corner. Pelvic anterior margin 0.9 times base length. In males, pelvic inner margins fused together forming a pelvic apron, elongated posteriorly and covering claspers. In females, the pelvic fin apex located obviously posterior to middle of fin, and angle of pelvic fin inner corner 63º–75º in paratypes.
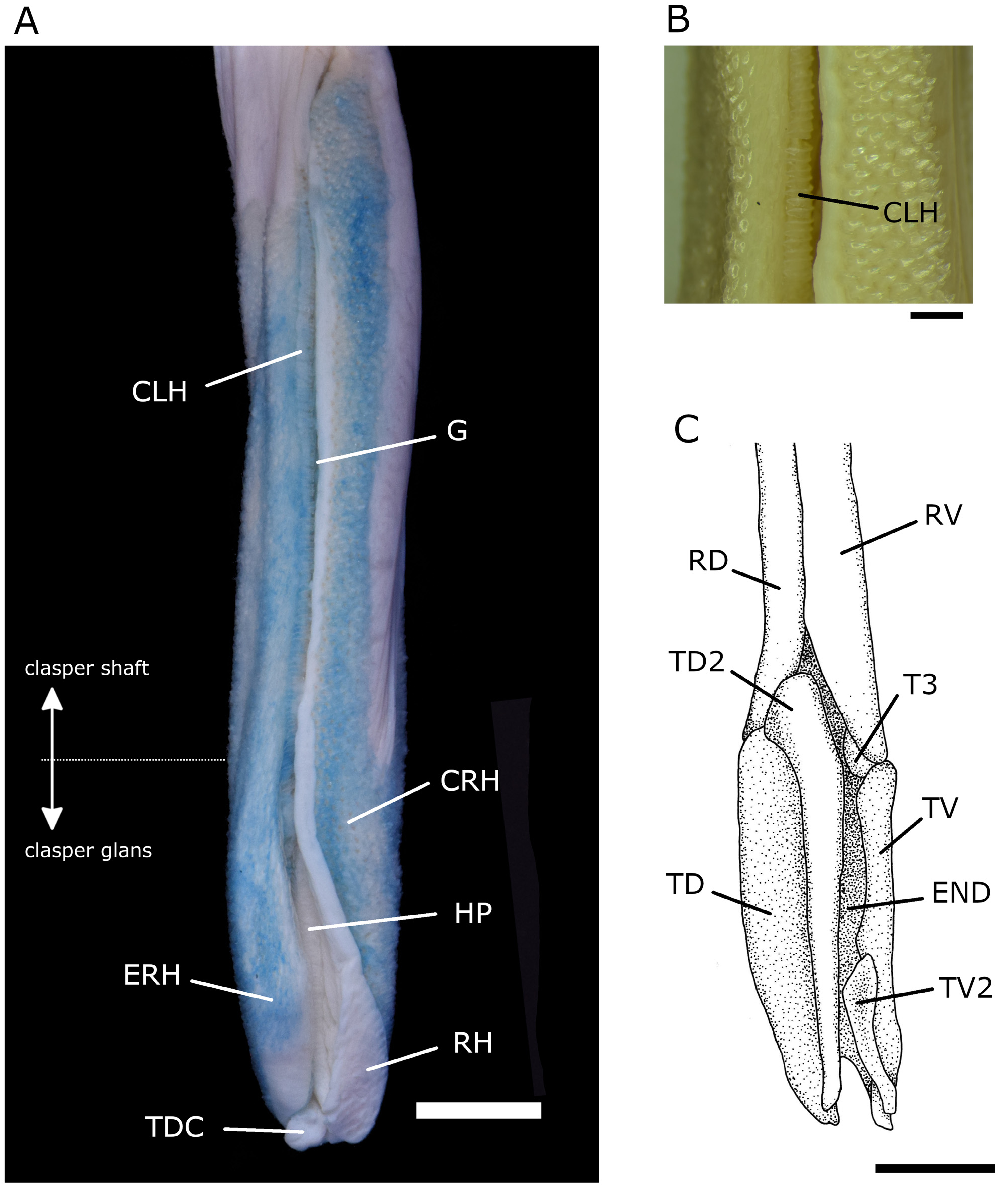
Claspers ( Fig. 5A View FIG ) moderately long and cylindrical, extending beyond free rear tips of pelvic fins in adult specimens. Clasper inner length 14.1 %TL (14.1–17.2 %TL in mature males of paratypes), 1.4 times (1.4–1.8 times in mature males of paratypes) pelvic anterior margin and 1.6 times (1.4–1.7 times) clasper outer length. Most of clasper surface (except part of medial surface of clasper shaft and part of exorhipidion, rhipidion, and terminal dermal cover) covered by dermal denticles with anteriorly directed crowns. Clasper hooks present, forming row from behind apopyle to posterior tip of exorhipidion, running along medial border of exorhipidion ( Fig. 5A, B View FIG ). Rhipidion well developed, partly covered medially by prominent exorhipidion and anteriorly by cover rhipidion. Rhipidion insertion point located in anterior portion of dorsal terminal 2 cartilage and extending to end of glans. Cover rhipidion expanded medially, reaching exorhipidion with nearly straight border at the shaft and meandering border at glans. Both cover rhipidion and exorhipidion cover clasper groove. Envelope, pseudopera, and pseudosiphon absent. Terminal dermal cover smooth, and contacting exorhipidion and rhipidion.
Clasper skeleton relatively simple ( Fig. 5C View FIG ). Ventral and dorsal terminal cartilage forms spoon-shaped structure; dorsal terminal begins anteriorly but ends together with ventral cartilage. Accessory terminal cartilage quadrangle situated posteriorly and inside ventral marginal cartilage. Dorsal terminal 2 cartilage beginning posteriorly and ventrally to dorsal marginal cartilage, elongated and rod-like, laterally positioned on dorsal terminal cartilage and extending throughout its length. Ventral terminal 2 cartilage elongated and oval, laterally and posteriorly positioned on ventral terminal cartilage and extending to half its length.
First dorsal fin ( Fig. 1 View FIG ) sub-rectangular and triangular, not falcate, with nearly straight anterior margin, round apex, and angular free rear tip. First dorsal base 0.8 times the interdorsal space. First dorsal anterior margin length 1.5 times its base, and first dorsal height 0.9 times its base.
Second dorsal fin ( Fig. 1 View FIG ) triangular, not falcate and smaller than first dorsal fin, with nearly straight anterior margin, straight posterior margin, rounded apex, and sharper free rear tip than first dorsal fin. Second dorsal fin base 1.4 times its height and 0.9 times dorsal–caudal distance. Second dorsal anterior margin 1.3 times its base. Second dorsal fin base 0.8 times first dorsal fin base, second dorsal fin height 0.6 times first dorsal fin height.
Anal fin ( Fig. 1 View FIG ) triangular, slightly high, apically narrow, and not falcate. Anal fin anterior margin nearly straight, apex rounded, free rear tip acutely pointed, and posterior margin straight. Anal fin base 0.8 times interdorsal space and 1.3 times dorsal–caudal distance. Anal fin anterior margin 1.5 times its posterior margin and 1.0 times its base. Anal fin height 0.5 times its base and 1.4 times caudal peduncle height. Anal fin larger than second dorsal fin, base 1.4 times second dorsal fin base, and anal fin height 1.0 times second dorsal fin height.
Caudal fin ( Fig. 1 View FIG ) narrow-lobed and asymmetrical, with developed terminal lobe. All caudal fin margins nearly straight, all tips rounded. Dorsal caudal margin without lateral undulations and crest of denticles. Dorsal caudal margin 0.3 times precaudal length and 2.1 times preventral caudal margin. Subterminal margin 0.6 times terminal margin.
Total vertebral counts 112 (109–115 in paratypes), monospondylous vertebral counts 34 (34–36), and precaudal vertebral counts 77 (71–78).
Intestinal valve type conicospiral, with 8 turns (7–8 turns in paratypes).
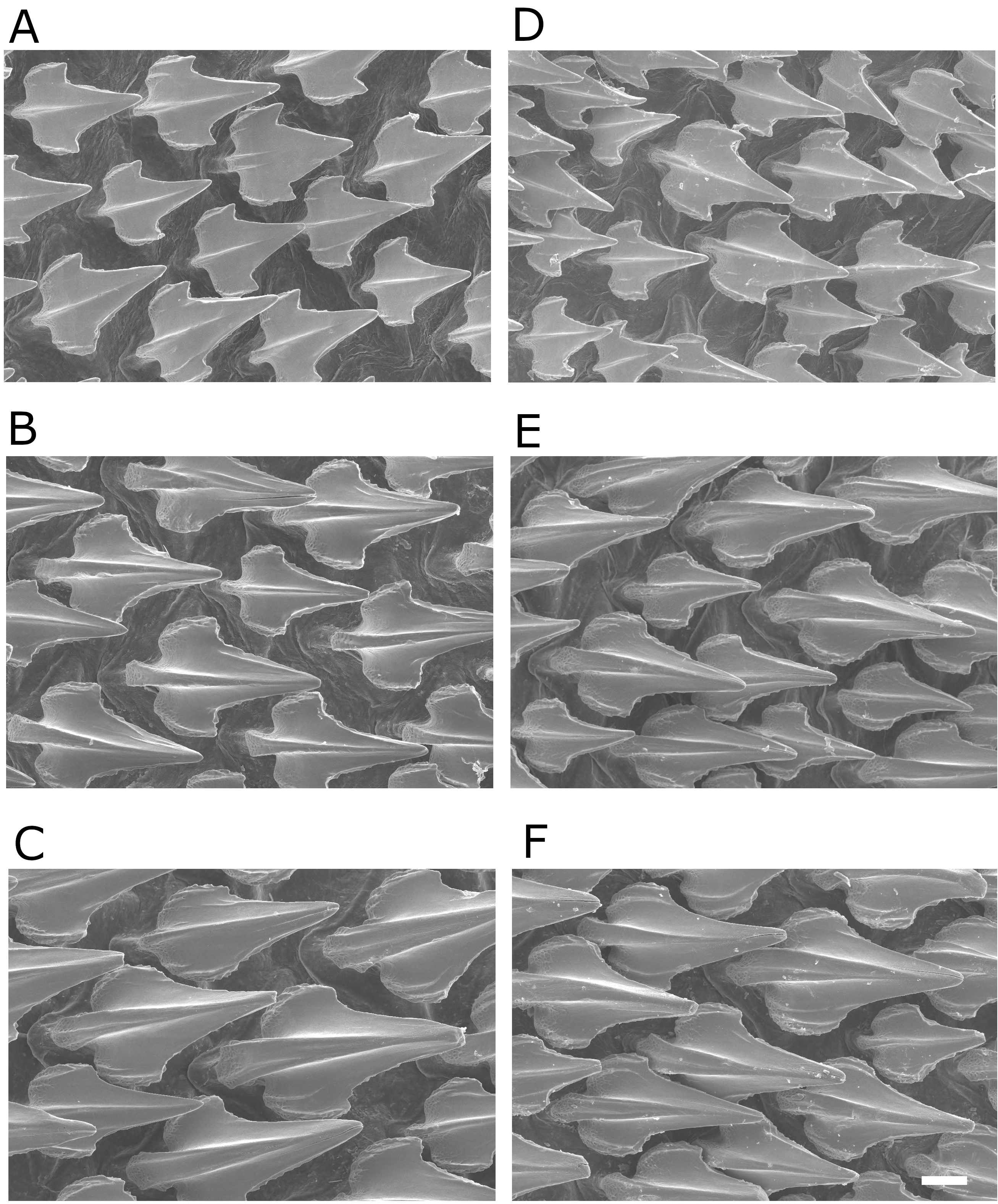
Dermal denticles ( Fig. 6 View FIG ) with flat, elongated, teardrop-shaped crowns. Crown with a strong medial ridge extending its entire length onto a long principal cusp. Dermal denticles above pectoral fin present three ridges, with medial ridge less prominent than in denticles of other regions. Denticles below dorsal fins are longer and present a prominent medial ridge, extending to distal tip of cusplet. In all regions, lateral ridges do not extend beyond intersection between principal cusp and cusplets, and are less prominent. Lateral cusps of denticles well developed above pectoral fin, but not well developed below dorsal fins.
Color pattern ( Figs. 1 View FIG , 4A, B View FIG ) with dark brown saddles on a brown background. Light spots present on dorsolateral surfaces and fins, spiracle-sized or larger. Dark spots present on lateral body and fins, smaller than spiracles and light spots. Outlines of dark spots not distinct. Light spots on saddles slightly smaller than on body, with spots largest on fins and smallest on saddles. Saddles darker than background, with pectoral saddles extending diagonally forward. Ventral region white to cream, turning dark anterior of the nostrils.
Distribution.
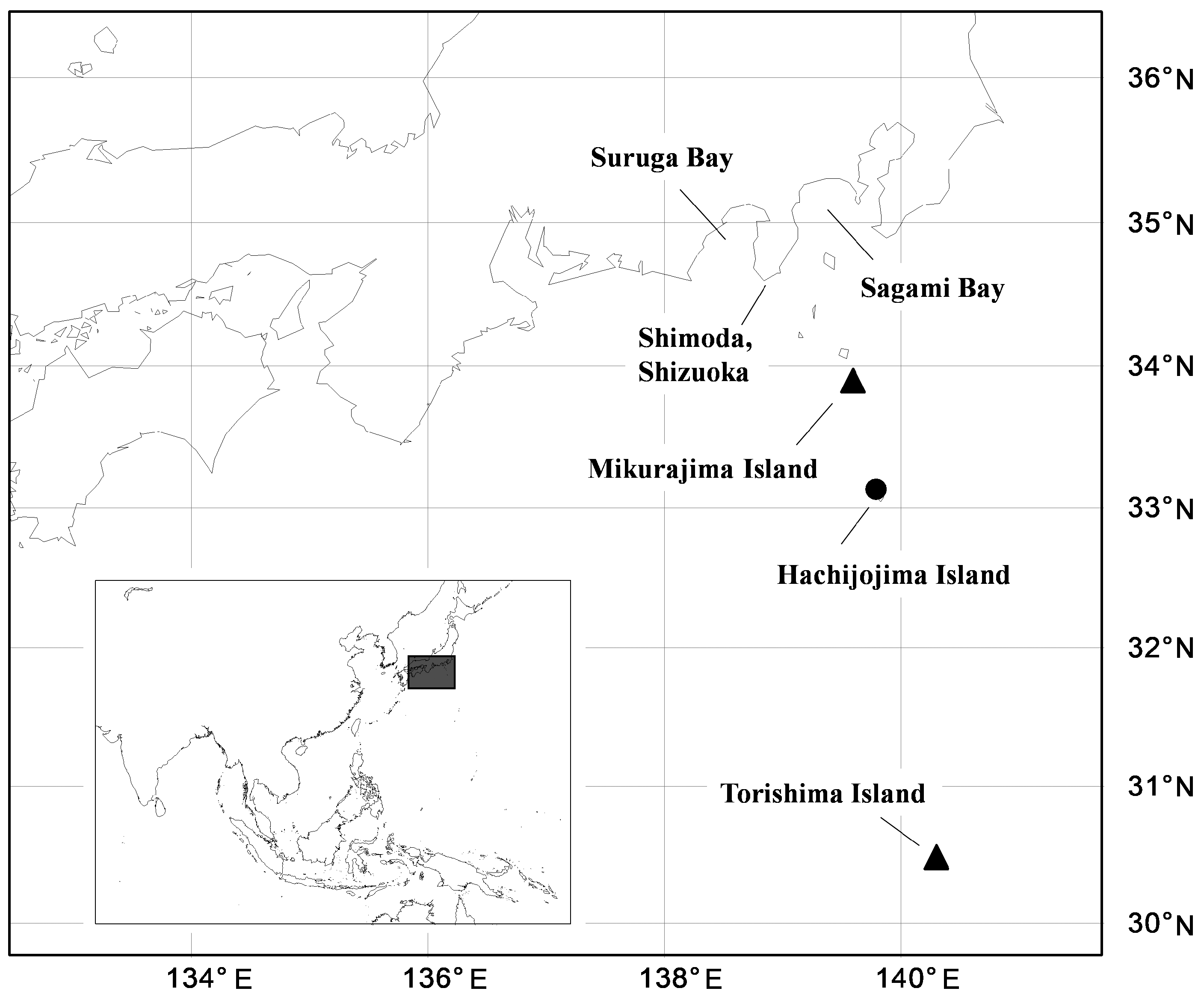
This species was recorded from the waters around the Izu Islands, Japan ( Fig. 7 View FIG ). All specimens were captured by longline fishing for Splendid alfonsino, at depths of ca. 100–200 m around Mikurajima Island, ca. 200–400 m around Hachijojima Island, and ca. 500–600 m around Torishima Island.
Biological data.
Males ranged in size from 294 to 416 mm TL (n = 6) and females from 301 to 386 mm TL (n = 22). Size at first maturity is not precisely defined, but males had well-developed claspers at 346 mm TL and females had egg cases at 342 mm. Reproduction is by single oviparity, one egg case for each oviduct. Egg cases are amber in color, with tendrils and surface irregularities that resemble wrinkles ( Fig. 8 View FIG ). Maximum case length without tendrils (ML) was 44.6–56.25 mm, and maximum case width was 19.45–22.5 mm (33.4–42.1 %ML) (n = 7).
Stomachs were found to contain small crustaceans, small bony fishes, and polychaets.
Genetic data.
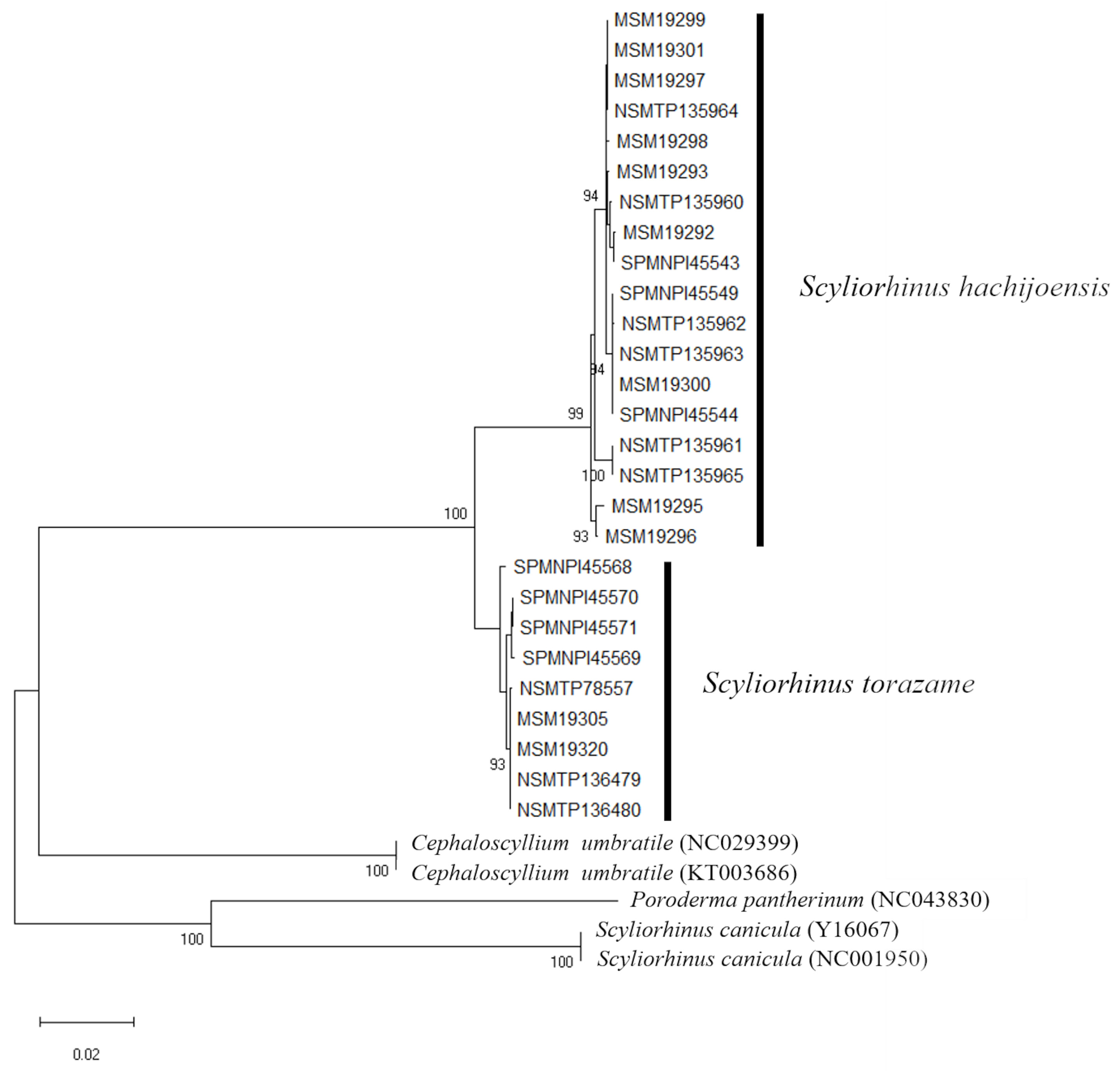
A total of 2164 base pairs (bp) from three mitochondrial DNA regions were examined: 487 bp for the 16S region, 664 bp for COI, and 1013 bp for Cytb. The monophyly of S. hachijoensis was strongly supported by molecular phylogenetic analysis ( Fig. 9 View FIG ). The genetic distances between S. hachijoensis and S. torazame were 0.3702 (16S), 0.0295 (COI) and 0.0403 (Cytb), respectively.
Etymology.
The species name “ hachijoensis ” refers to the species’ main collection area, Hachijojima Island. The English name is derived from “Cinderella”, because the dark spots on the body surface are similar to black ashes “cinder”. The Japanese name “Fukami” means “deep sea”.
Remarks.
Clasper hooks are a unique character observed in S. torazame ( Soares & de Carvalho 2019, 2020), and the same character was also observed in S. hachijoensis .
Both male and female S. hachijoensis can be easily distinguished from eleven other Scyliorhinus species by color pattern: S. cervigoni Maurin & Bonnet, 1970 , S. garmani (Fowler, 1934) , S. meadi Springer, 1966 and S. retifer (Garman, 1881) have no light spots, S. capensis (Müller & Henle, 1838) has light yellow to golden spots, and S. torrei Howell-Rivero, 1936 has beige to cream spots. Scyliorhinus canicula , S. cabofriensis Soares, Gomes & de Carvalho, 2016 , S. cervigoni , S. garmani , S. haeckelii (Miranda Ribeiro, 1907) and S. stellaris (Linnaeus, 1758) have conspicuous dark spots, and S. boa (Goobe & Bean, 1896) has various dark spots ( Springer 1979; Ebert et al. 2013a; Soares & de Carvalho 2019). Scyliorhinus comoroensis Compagno, 1988 and S. duhamelii (Garman, 1913) (as well as S. canicula , S. cervigoni , S. garmani and S. stellaris , which are also listed above) can be distinguished from S. hachijoensis because the nasal flaps reach or cover the upper lip ( Springer 1979; Ebert et al. 2013a; Soares & de Carvalho 2019). Scyliorhinus hesperius Springer, 1966 and S. ugoi Soares, Gomes & Gadig, 2015 are very similar to S. hachijoensis in color pattern, but differ in the number of monospondylous vertebrae and size of maturity. The number of monospondylous vertebrae is 39–42 in S. hesperius and 38–39 in S. ugoi . Adult males mature at least 420 mm TL in S. hesperius , and by at least 450 mm TL and 470 mm TL in male and female specimens of S. ugoi , respectively. By contrast, the number of monospondylous vertebrae in S. hachijoensis is 34–36 and the size at maturity is less than 350 mm TL. In addition, S. ugoi has saddles with antero- and posteromedial projections ( Springer 1979; Ebert et al. 2013a; Soares et al. 2015; Soares & de Carvalho 2019).
Scyliorhinus torazame is similar in external characters to S. hachijoensis , but differs in fin height and fin angle. In S. torazame , anal fin height is less than 4 %TL and approximately equal to caudal peduncle height (as compared to more than 4 %TL and higher than the caudal peduncle in S. hachijoensis ) ( Table 1 View TABLE 1 ), the pectoral fin angle in adults is 70º–96º and the pelvic fin angle in adult females is 40º–65º (as compared to a pectoral fin angle of 90º–102º and pelvic fin angle of 63º–75º in S. hachijoensis ), the pelvic fin apex is located in the middle of the fin (vs. apex obviously behind the middle of the fin in S. hachijoensis ) ( Figs. 1 View FIG , 4 View FIG ), and the dermal denticles present five ridges (vs. three ridges in S. hachijoensis ). Also, S. torazame has no dark spots. Finally, whereas the egg case surface is irregularities that resemble wrinkles in S. hachijoensis , the egg case is smooth in S. torazame ( Fig. 8 View FIG ).
Previously, S. torazame had not been reported from any waters farther south than Taiwan. Also, reports of this species from Okinawa have been rare ( Springer 1979; Carpenter 1998; Ebert et al. 2013a; Soares & de Carvalho 2019). In this study, we identified numerous specimens from the Ryukyu Islands (specifically, the islands of Okinawa and Haterumajima) and Taiwan, including the first confirmed specimen from the South China Sea, thus expanding the known distribution of S. torazame (see APPENDIX 1).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Scyliorhinus hachijoensis
| Ito, Nanami, Fujii, Miho, Nohara, Kenji & Tanaka, Sho 2022 |
Scyliorhinus sp.
| Hagiwara, S. 1993: 1 |