
Scyliorhinus torazame ( Tanaka, 1908 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4601.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:8A695352-8382-458F-A86A-17A198F780CA |
|
persistent identifier |
https://treatment.plazi.org/id/03B94378-D00A-0672-FF7D-FDD7FB31AB82 |
|
treatment provided by |
Plazi |
|
scientific name |
Scyliorhinus torazame ( Tanaka, 1908 ) |
| status |
|
Scyliorhinus torazame ( Tanaka, 1908) View in CoL
( Figs. 77–85 View FIGURE 77 View FIGURE 78 View FIGURE 79 View FIGURE 80 View FIGURE 81 View FIGURE 82 View FIGURE 83 View FIGURE 84 View FIGURE 85 , Tabs. 3 View TABLE 3 , 17 View TABLE 17 , 18 View TABLE 18 )
Common names: torazame catshark ( Japan).
Catulus torazame Tanaka, 1908: 6 , pl. 2, figs. 2a, 2b (original description, type locality: Misaki, Japan); Garman, 1913: 77 (brief account, classification); Tanaka, 1931: 5, fig. 8 (catalogued); Matsubara, 1936: 36, fig. 26 (compilation); White, 1937: 107, 117 (listed, systematics); Kamohara, 1950: 4 (listed, Japan); Honma, 1952: 139 (listed).
Scyliorhinus rudis Pietschmann, 1906: 2 View in CoL (original description, type locality: Japan); Pietschmann, 1908: 699 –707, pl. 1, figs. 9–14 (brief account, Japan); Regan, 1908: 457 –458 (listed, classification) [synonymy follows Compagno, 1984: 367 –368].
Halaelurus rudis: Tanaka, 1911: 13 , pl. 3, fig. 12 [synonymy follows Jordan, Tanaka & Snyder, 1913: 10].
Halaelurus torazame : Jordan, Tanaka & Snyder, 1913: 10 (catalogue, Japan); Schmidt, 1930: 48 (clasper description).
Scyliorhinus torazame: Fowler, 1941: 36 View in CoL (catalogue, Philippines); Mori, 1952: 18 (listed, Japan); Herre, 1953: 12 (listed, Philippines); Matsubara, 1955: 108, pl. 3, fig. 9 (compilation); Mori, 1956: 2 (listed, Japan); Chu & Wang, 1964: 675 (listed, Japan); Lindberg & Legeza, 1967: 41, fig. 25 (catalogue, southern Japan); Ueno, 1971: 68 (listed); Nakaya, 1975: 15 –22, figs. 4–9 (brief account, systematics); Springer, 1979: 144 –146, figs. 95, 96 (taxonomic review); Compagno, 1984: 367 –368 (FAO catalogue); Masuda et al., 1984: 4, pl. 3B (brief account); Nakaya & Shirai, 1992: 40, 44 (biogeography); Carpenter & Niem, 1998: 1205 (FAO catalogue, Western Central Pacific); Compagno, 1999: 480 (listed); Randall & Lim, 2000: 579 (listed, southern China Sea); Compagno et al., 2005: 253, pl. 42 (compilation); Ebert et al., 2013a: 374, 384, pl. 52 (compilation); Sinohara et al., 2014: 232 (listed, Japan); Weigmann, 2016: 44 (listed).
Scyliorhinus tokubee Shirai, Hagiwara & Nakaya, 1992: 9 View in CoL –16, figs. 1–3 (original description, type locality: Izu Peninsula, southern Japan); Compagno et al., 2005: 253, pl. 42 (compilation); Ebert et al., 2013a: 374, 383, pl. 52 (compilation); Ebert et al., 2013b: 320 (listed, Taiwan); Weigmann, 2016: 44 (listed) [new synonym].
Holotype. ZUMT 953 View Materials , male, 450 mm TL ( Misaki , Sagami, Japan). Status: lost.
Additional material examined. 131 specimens (see Appendix).
Diagnosis. Scyliorhinus torazame differs from all congeners by presenting specialized hooks in the clasper (vs. absent in all other species); pelvic apron extending through almost the entire length of pelvic inner margins (vs. extending to 2/3 length of pelvic inner margins in S. boa , S. cabofriensis , S. cervigoni , S. comoroensis , S. haeckelii , S. hesperius , S. meadi , S. retifer , S. stellaris , and S. ugoi ); males presenting distal tips of pelvic fins straight (vs. tapered in the other species); body beige to light brown with cream to beige spots (vs. light spots absent in S. cervigoni , S. garmani , S. meadi , and S. retifer ; yellow to golden spots in S. capensis ), and spots predominantly spiracle-sized (vs. predominantly smaller than spiracles in S. boa , S. cabofriensis , S. canicula , S. duhamelii , S. stellaris , and S. ugoi ). The following combination of characters, although less conspicuous, also helps distinguish this species: saddles darker than the background color (vs. inconspicuous in S. boa , S. cabofriensis , S. cervigoni , S. garmani , and S. torrei ); anterior nasal flaps not reaching upper lip (vs. flaps reaching upper lip, and sometimes covering it, in S. canicula , S. cervigoni , comoroensis , S. duhamelii , S. garmani , and S. stellaris ); interdorsal distance 0.6–1.0 times smaller than anal base (vs. greater than anal base in S. boa , S. cabofriensis , S. haeckelii , S. hesperius , S. meadi , S. retifer , S. torrei , and S. ugoi ); mandibular canal of lateral line system with 5 to 6 pores (vs. 3–4 in S. hesperius ); oral canal of lateral line system with 7–10 pores (vs. 10–12 in S. duhamelii ; 5–6 in S. hesperius ; 9–13 in S. torrei ); commissural teeth with one cusplet and principal cusplet laterally situated (vs. two or more in other species, except in S. cervigoni , S. meadi and S. torrei ); clasper with terminal dermal cover smooth (vs. rough in S. canicula and S. capensis ); cover rhipidion covered by dermal denticles (vs. uncovered in S. boa , S. cervigoni and S. retifer ); terminal 3 cartilage present (vs. absent in S. cabofriensis , S. cervigoni , S. comoroensis , S. duhamelii , S. haeckelii , S. stellaris , S. torrei , and S. ugoi ); dorsal terminal 2 cartilage elongated and similar in size to dorsal terminal cartilage (vs. reduced and subtriangular in S. cabofriensis , S. capensis , S. cervigoni , S. haeckelii , and S. ugoi ; dorsal terminal 2 less than half length of dorsal terminal cartilage in S. boa , S. canicula , S. duhamelii , S. retifer , S. stellaris , and S. torrei ); counts of monospondylous vertebrae 32–37 (vs. counts lower in other species, except in S. canicula , S. duhamelii , S. haeckelii , and S. torrei ); adult males between 360–400 mm TL and adult females between 370–420 mm TL (vs. adult reach greater sizes at sexual maturity in S. capensis , S. cervigoni , S. meadi , S. stellaris , and S. ugoi ).
Description. Morphometric and meristic data are given in Table 17 View TABLE 17 , and neurocranial measurements in Table 18 View TABLE 18 .
Body robust and cylindrical, tapering considerably posterior to cloaca ( Fig. 77 View FIGURE 77 ). Prepectoral length 0.4 times the prepelvic length. Trunk shorter than tail; snout-vent length 0.8 times vent-caudal length. Pectoral-pelvic space 1.6–1.7 times the pelvic-anal space. Interdorsal space 1.8–2.9 times the dorsal-caudal space ( Tab. 17 View TABLE 17 ). No interdorsal, postdorsal or postanal ridges; lateral crest on caudal peduncle absent.
Head moderately broad and depressed; head length 1.7 times head width ( Fig. 77 View FIGURE 77 ). Snout short, preoral length 0.4–0.6 times mouth width and 0.6–0.7 times smaller than preorbital length. Prenasal length 0.7 times internarial space; preorbital length 0.3–0.6 times interorbital space.
Eye large and slitlike, eye length 2.4–4 times its height and 0.2–0.3 times smaller than head length ( Fig. 77 View FIGURE 77 ). Eye dorsolateral on head, with lower edge medial to horizontal head rim in dorsal view; subocular ridge strong. Nictitating lower eyelid of rudimentary type, with shallow subocular pouch and secondary lower eyelid free from upper eyelid. Spiracle close behind but well separated from eyes, dorsolaterally on head and somewhat lower than level of eye notch. Spiracle diameter goes 3.5–4.2 times in eye length and 6.7–11.5 times in interorbital width.
First two gill openings about equally wide; first one twice as long as fifth. All gill openings slightly concave and not elevated on dorsolateral surface of head; gill filaments not visible externally.
Nostril with broad incurrent aperture, without nasoral groove or nasal barbel, and small and oval excurrent aperture. Anterior nasal flap large, triangular, and covering posterior nasal flap and excurrent aperture, extending just anterior to mouth, close to the upper lip but not touching it ( Figs. 78 View FIGURE 78 A–B). Mesonarial ridge distinct but not exceeding the posterior border of the anterior nasal flap. Posterior nasal flap rectangular, situated on the posterior border of the excurrent aperture. Mesonarial superior and inferior flaps conical and corresponding to 1/4 of anterior nasal flap. Internarial space 0.7–0.9 times smaller than interorbital space.
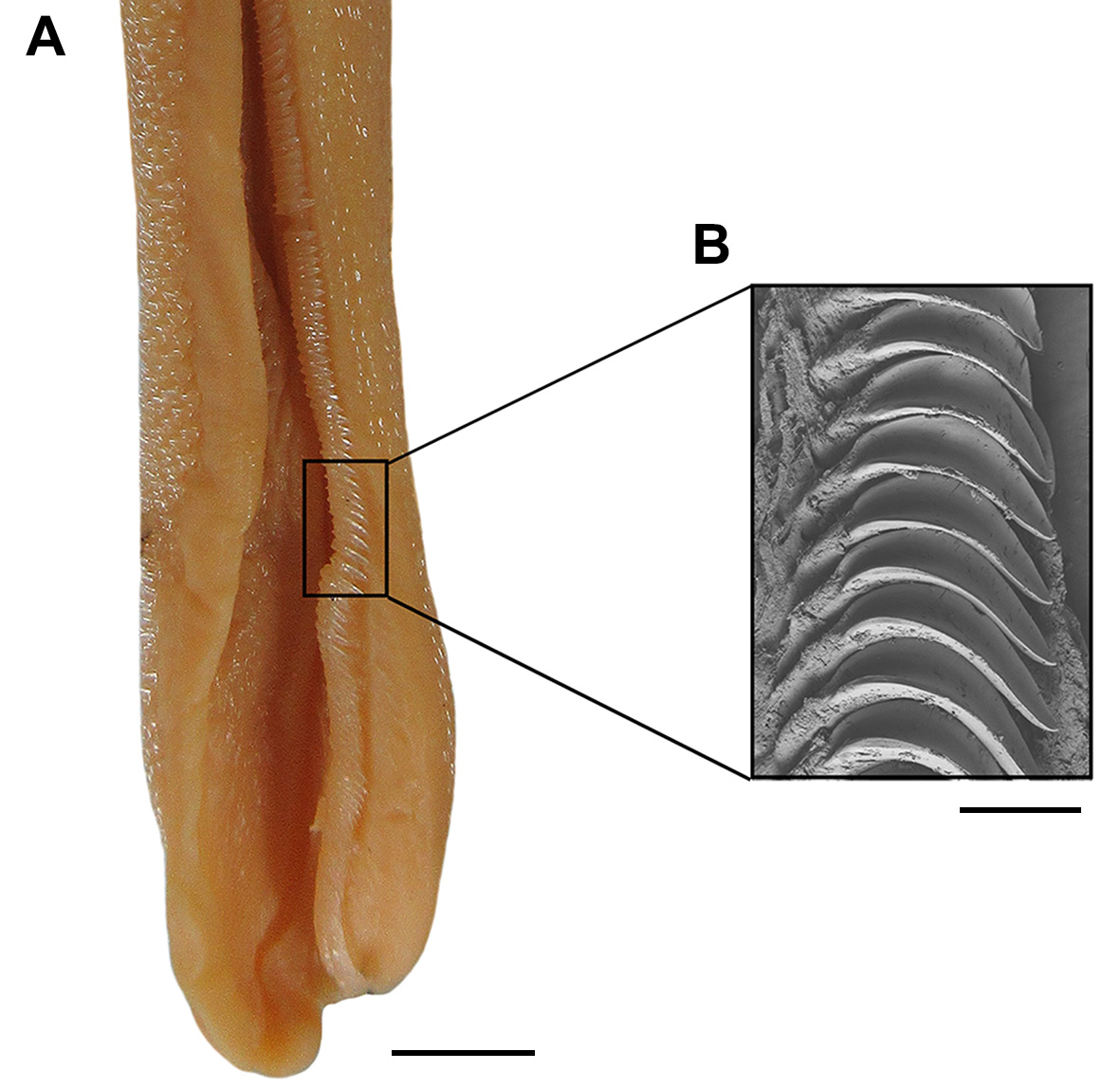
Mouth arched, moderately wide and short, its length goes 1.8–2.0 times in mouth width ( Figs. 78 View FIGURE 78 A–B). Lower labial furrow short and narrow, 3.9–6.4 times smaller than mouth width. Dorsal labial cartilage 1.3 times the ventral cartilage; anterior tip of dorsal labial cartilage reaching the orbital process of the palatoquadrate. Tongue flat and rounded, light-colored, with oral papillae hardly detectable.
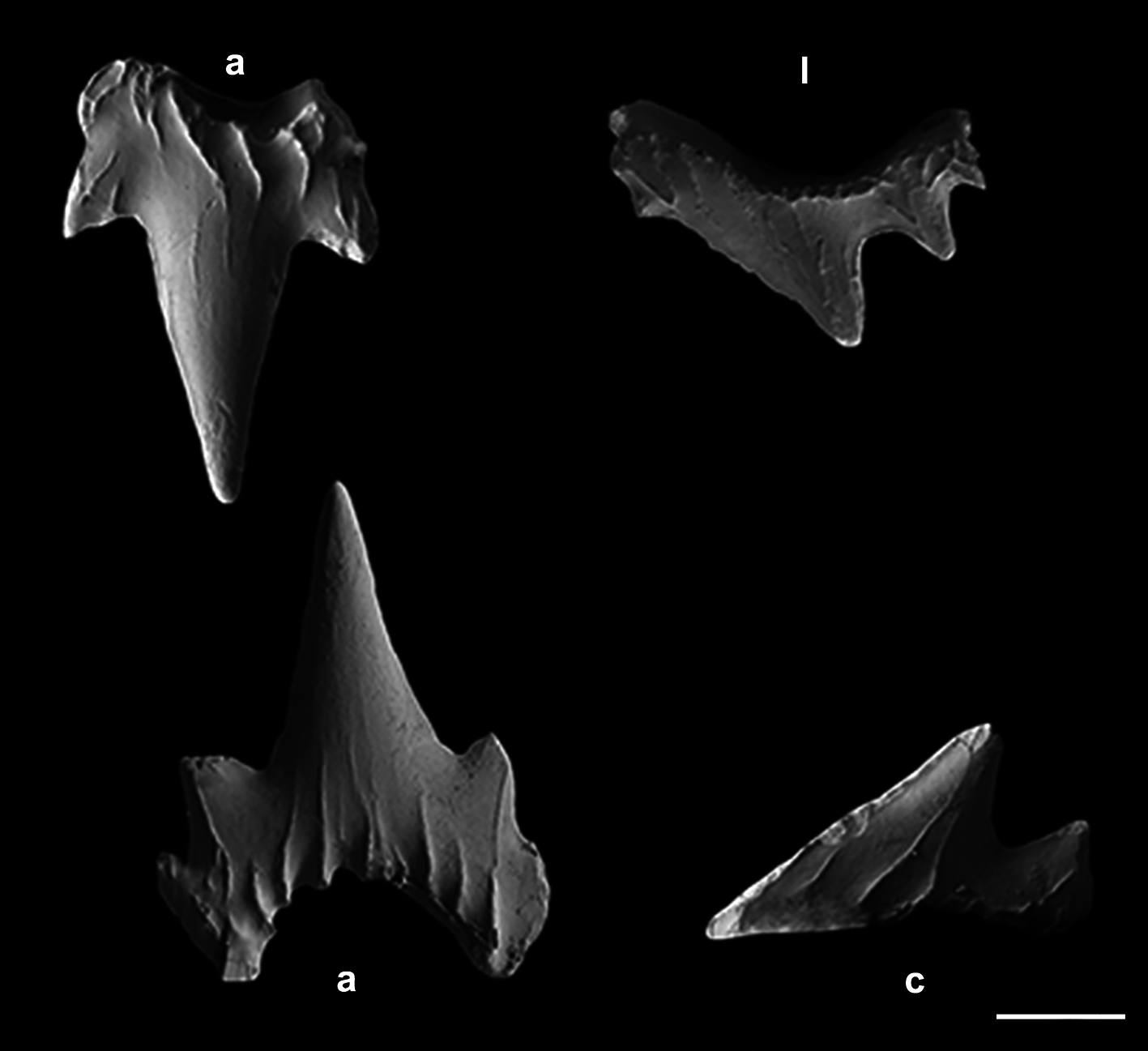
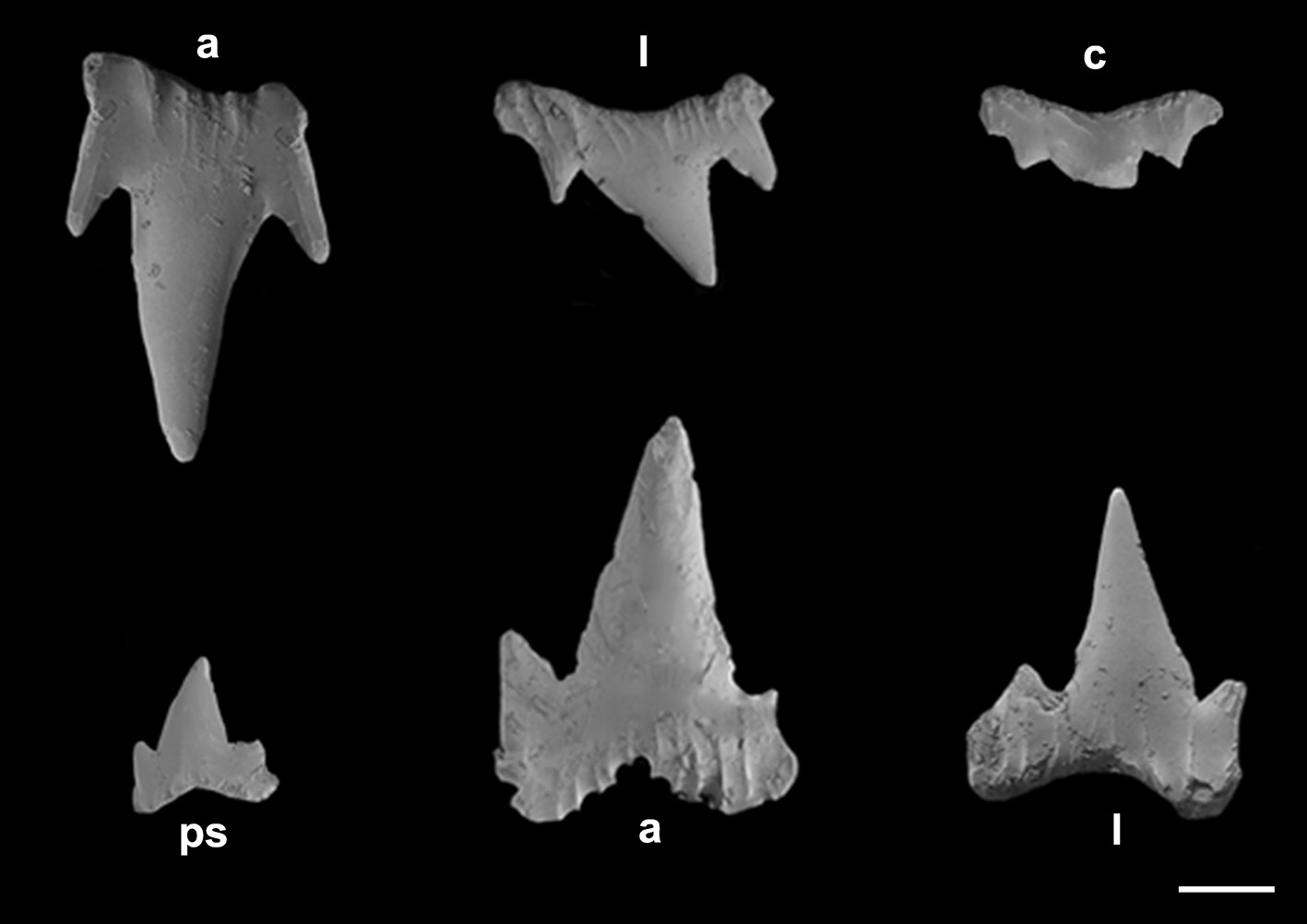
Monognathic heterodonty gradual well developed; anterior teeth abruptly larger than the parasymphysial ones and lateral teeth smaller distally, with smaller and thicker principal cusps. Sexual heterodonty well pronounced with females presenting anterior teeth with marginal cusplet well-developed and proximal one more tapered and oblique in males ( Figs. 79 View FIGURE 79 , 80 View FIGURE 80 ). Tooth counts 24–38 26–38/22–42 23–40. Parasymphysial teeth with a principal cusp flanked by one cusplet on each side; cusplets half the height and the width of the principal cusp. Protuberances on medial portion of the crown base and striae restricted to the crown base. Anterior teeth larger than the parasymphysial and principal cusp less stout. Anterior teeth with two to four cusplets; marginal cusplets smaller and half the height or less of the principal cusp in upper teeth and more than half the height in lower ones. Protuberances on the crown base; striae restricted to the crown base in lower teeth and extending to half the height of the principal cusp in upper ones. Lateral with three or four cusplets; marginal cusplets half the height of proximal ones. Upper teeth with principal cusp slightly oblique; nearly straight in lower ones. Protuberances on the crown base; striae restricted to the crown base in lower teeth and extending to half the height of the principal cusp in upper ones. Commissural teeth with one cusplet; principal cusp stouter and laterally situated. Cusplet corresponding to 2/3 the height and the width of the principal cusp. Protuberances on the crown base and striae throughout the principal cusp. Ectodermal pits inconspicuous or restricted to the crown base in lateral and commissural teeth.
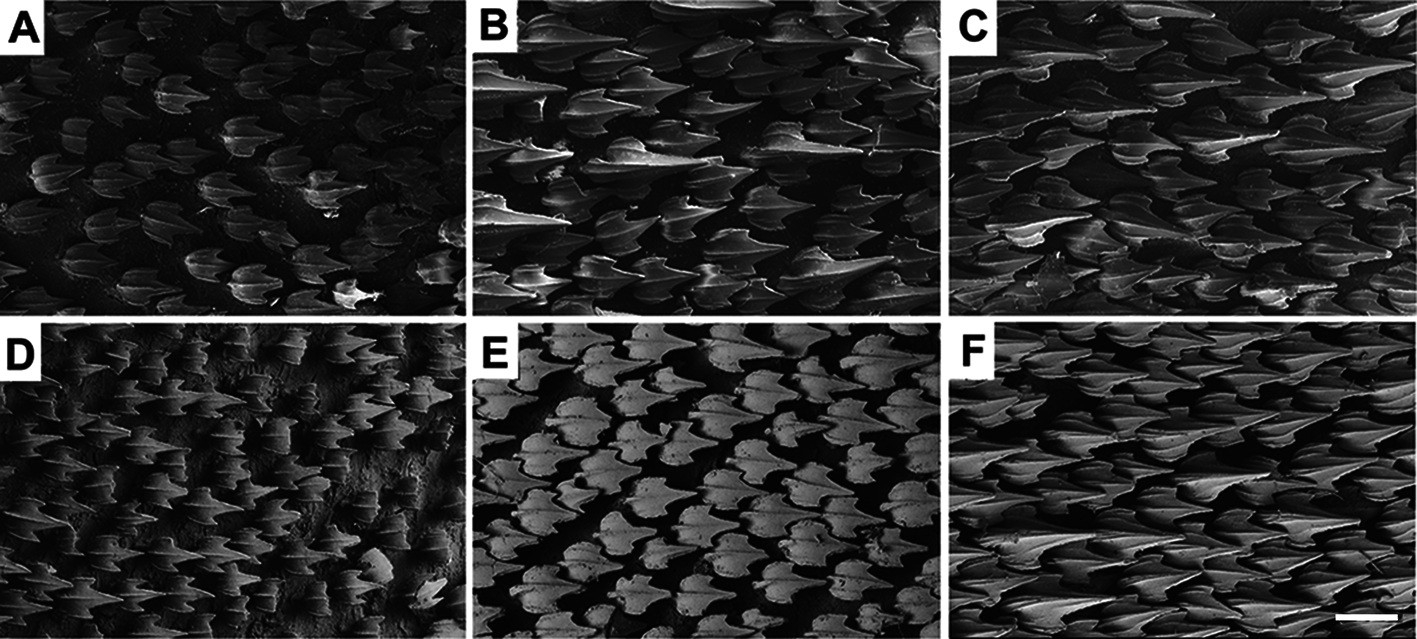
Lateral trunk denticles with flat, elongated teardrop-shaped crowns, 1.3–1.8 times as long as wide ( Tab. 3 View TABLE 3 ); anterior part covered with ectodermal pits. Crown with a strong medial ridge extending its entire length onto long principal cusp. Dermal denticles above the pectoral fin presenting five ridges, medial ridge less prominent than in denticles of other regions and lateral ridges not extending beyond the intersection between principal cusp and cusplets. Denticles below dorsal fins longer and presenting median and lateral ridges prominent, extending to distal tip of cusplets ( Fig. 81 View FIGURE 81 ).
Pectoral base 0.7–0.8 times mouth width ( Fig. 78C View FIGURE 78 ). Pectoral anterior margin 2–2.3 times its base and 1.4–1.6 times the posterior margin. Pectoral fin skeleton aplesodic with radials mostly divided into three segments. Propterygium and mesopterygium trapezoidal; the former smaller than the latter. Propterygium with one proximal segment; mesopterygium with 3–4 proximal segments fused proximally. Metapterygium with 9 segments. Metapterygial axis rectangular and corresponding to 1/5 of metapterygium.
Pelvic fin subrectangular in males and trapezoidal in females ( Fig. 78F View FIGURE 78 ); pelvic anterior margin 0.8–0.9 times the posterior margins and 1.4–2.4 times the pelvic base. Pelvic inner margins of males fused by almost all their extension, with rectangular edges; claspers of juveniles covered by the pelvic apron and evident only with the lifted apron.
Clasper moderately long and cylindrical ( Fig. 78F View FIGURE 78 ), extending beyond free rear tips of pelvic fins in adult specimens; clasper inner length 0.8–1.0 times the pelvic anterior margin, 1.4–2.4 times the clasper outer length and 5.4–5.6 times the clasper base. Most of clasper surface except dorsomedial surface of glans, medial border of cover rhipidion, rhipidion, and terminal dermal cover, covered by dermal denticles with anteriorly directed crowns ( Fig. 82A View FIGURE 82 ). Clasper hooks present, forming a row from the anterior 1/4 of marginal cartilages to the posterior tip of claspers glans, running along the medial border of exorhipidion and covered by it posteriorly ( Fig. 81 View FIGURE 81 ). Rhipidion well-developed, partly covered medially by a prominent exorhipidion and anteriorly by the cover rhipidion; insertion of rhipidion at anterior portion of dorsal terminal 2 cartilage and extending to the end of glans. Cover rhipidion expanded medially reaching an exorhipidion with nearly straight border; both cover rhipidion and exorhipidion covering the clasper groove. Envelope absent; pseudosiphon distinct and robust. Terminal dermal cover smooth and extending for 1/3 of ventral terminal cartilage, contacting the exorhipidion and cover rhipidion.
Clasper skeleton relatively simple ( Fig. 82B View FIGURE 82 ). Ventral and dorsal terminal cartilages forming a spoon-shaped structure; ventral terminal beginning anteriorly, but ending together with the dorsal cartilage. Terminal 3 cartilage rectangular, situated on anteromedial border of ventral terminal cartilage and ventrally to ventral terminal 2 cartilage. Dorsal terminal 2 cartilage elongated and rod-like, laterally positioned on the dorsal terminal cartilage and extending throughout the length of this. Ventral terminal 2 cartilage anteriorly expanded, tapering posteriorly and corresponding to 3/4 ventral terminal cartilage; ventral terminal 2 cartilage situated medially to ventral terminal and beginning at the intersection between ventral marginal and terminal cartilages.
First dorsal fin subrectangular and triangular, with nearly straight anterior margin, rounded apex and angular free rear tip ( Fig. 78 View FIGURE 78 ). First dorsal fin origin opposite or slightly posterior to the pelvic fin insertion. First dorsal fin insertion opposite to the anterior 1/3 of pelvic-anal distance. Anterior margin 1.4–1.5 times first dorsal fin base; first dorsal fin height 0.6–0.7 times its base.
Second dorsal fin triangular and smaller than the first ( Fig. 78 View FIGURE 78 ). Second dorsal fin origin opposite to posterior 2/5 of anal base and its insertion posterior to posterior tip of anal fin. Anterior margin 1.3–1.4 times base of second dorsal fin; second dorsal base 1.6–1.8 times its height and 1.3–1.7 times the dorsal-caudal distance. First dorsal fin 1.3 times larger than the second dorsal fin.
Anal fin triangular, apically narrow and not falcate ( Fig. 78 View FIGURE 78 ); anal fin base 1.6–1.7 times the second dorsal fin base. Anal anterior margin nearly straight, apex narrowly rounded, free rear tip acutely pointed, and inner margin straight. Anal fin base 0.9–1.1 times the interdorsal distance and 2–2.7 times the dorsal-caudal distance. Anal fin anterior margin 1.5–1.6 times the posterior margin; anal fin height 0.3–0.4 times its base.
Caudal fin narrow-lobed and asymmetrical ( Fig. 78 View FIGURE 78 ). Dorsal caudal lobe 1.7–1.8 times larger than preventral lobe; subterminal caudal margin as long as the terminal margin. Caudal crest of enlarged denticles absent on caudal fin margins.
Neurocranium broad and somewhat flattened, corresponding to 7.8–8.7% TL. Rostrum length 1.3–1.7 times the distance between lateral rostral cartilages; distance greater in males than in females. Nasal capsule wider than long, oval-shaped and expanded laterally; width 1–1.1 times its length. Width across nasal capsules proportionately narrower in males (73.3–77.3% NL) than in females (77.6–90.1% NL). Anterior fontanelle broad and subrectangular in males and wider and heart-shaped in females; epiphyseal notch very prominent. Basal plate flat with narrow borders, its width 1.8–2.3 times smaller than nasobasal length. Orbital region 2.2 times smaller than the nasobasal length. Otic capsule short, its length 2.9–4.1 times smaller than nasobasal length and width 2.2 times otic capsule length. Width across postorbital processes 1.2–1.3 times the preorbital processes width ( Tab. 18 View TABLE 18 ).
Coloration in alcohol. Body beige with seven to eight saddles darker than the background color. Spots cream colored, spiracle-sized or slightly greater; spots present or not on body and predominantly within saddles ( Fig. 78 View FIGURE 78 ). Some specimens presenting rosette-shaped spots forming aggregations ( Fig. 78C View FIGURE 78 ). Belly and ventral surface of paired and anal fins without spots, cream in color.
Distribution. This species is distributed along the continental shelves of eastern and western Japan, with the northernmost record for Otaru, Sapporo, Hokkaido island (43°N); known also from the eastern coast of South Korea, and with records from East China Sea and northeastern Taiwan, next to Yilan (24°40’N) ( Fig. 84 View FIGURE 84 ).
Biological data. Adult males between 360–400 mm TL; largest male specimen examined and maximum size reported 778 mm TL. Adult females between 370–420 mm TL; largest female specimen examined 478 mm TL. Egg capsules translucent yellow with smooth surface. Anterior tip slightly concave and posterior tip truncate, with long tendrils. Mean values for egg capsule dimensions: 55 mm length and 19 mm width (n = 3).
This species is known to enter shallow coastal waters ( Nakaya 1975; Yamada, 1986), with records from the Tsushima Strait (120 m in depth) and Korea Strait (140 m in depth) between the Okinawa Valley and Japan Sea. Conservation status ‘Least Concern’ ( Nakaya et al. 2009).
Etymology. The specific name ‘torazame’ refers to the common name attributed to this species in Japan, according to Tanaka (1908).
Remarks. Nakaya (1975), in his comprehensive study of Japanese catsharks, examined 24 specimens of S. torazame , including 11 males and 13 females, measuring between 245–500 mm TL. Shirai et al. (1992) described a new species, S. tokubee , and examined eight specimens of S. torazame (four males and four females) as comparative material. Among the characters indicated by these authors to distinguish S. tokubee from S. torazame were: mouth width at about two times smaller than head width, interdorsal distance slightly smaller than mouth width and equal to prespiracular length, tooth counts, and size of dermal denticles.
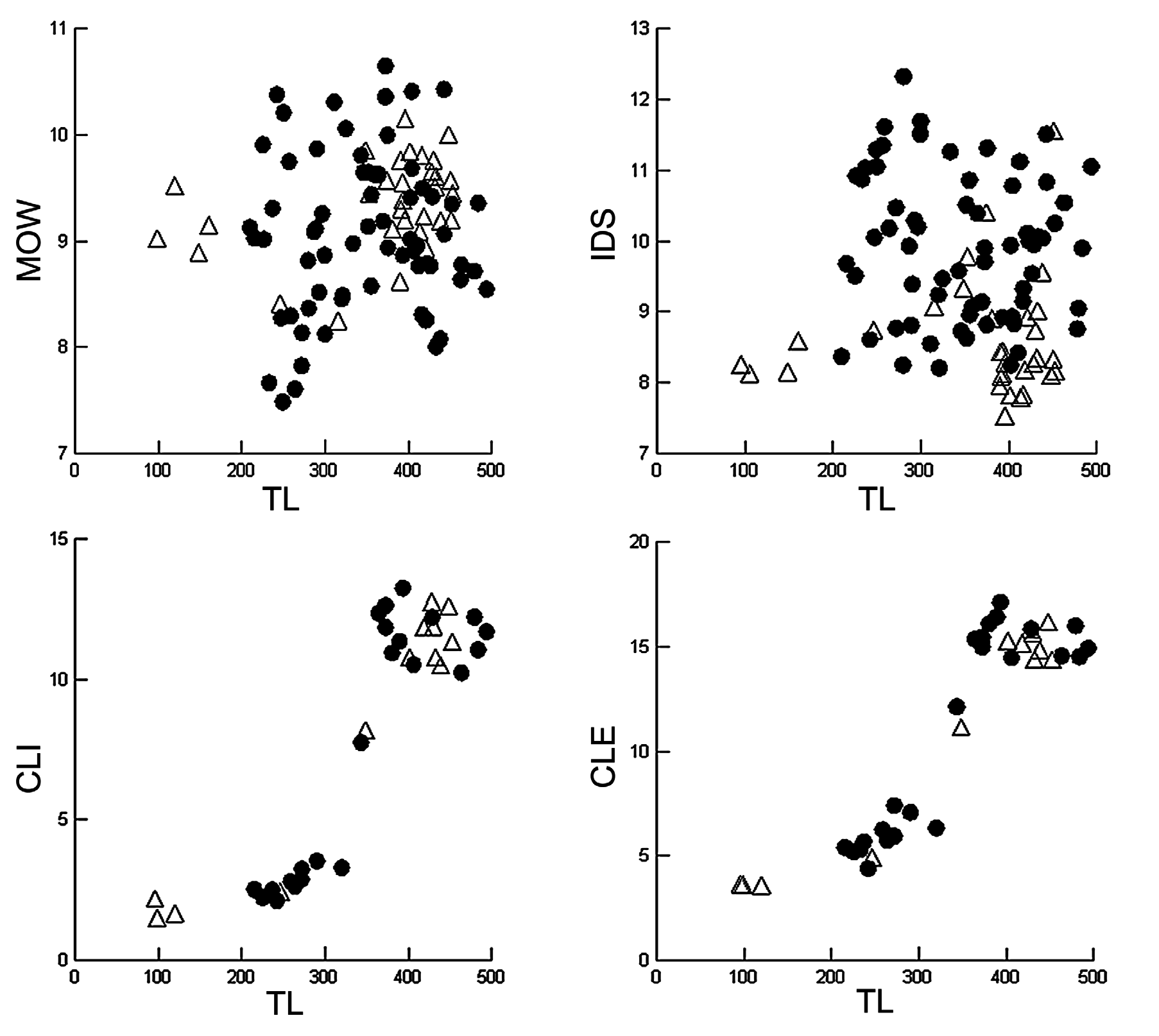
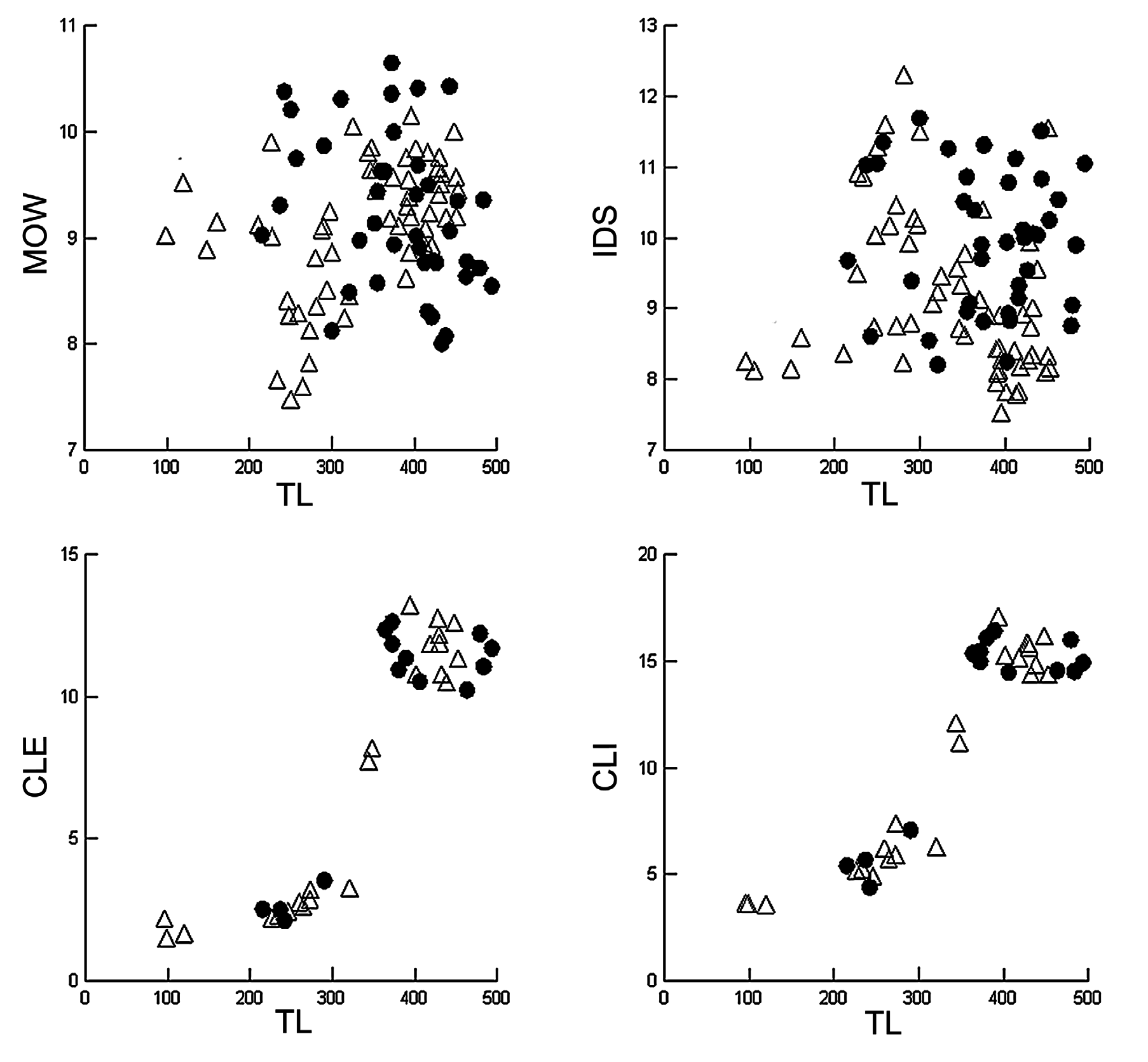
In the present study, 125 specimens of S. torazame and the type series of S. tokubee (n = 6), measuring between 95–778 mm TL, were examined. Specimens were separated according to color pattern (with/without light spots) and capture locality (eastern/western Japan). A graphic analysis of the measurements cited by Shirai et al. (1992) as diagnostic characters for S. tokubee (interdorsal distance and mouth width) was performed here ( Figs. 85 View FIGURE 85 , 86 View FIGURE 86 ). Additionally, clasper measurements were used to compare curves of sexual maturity ( Figs. 85 View FIGURE 85 , 86 View FIGURE 86 , C–D). According to the graphs, specimens captured close to Shizuoka (with spots) and Fukushima (without spots) present lower values for interdorsal distance around 8% TL ( Fig. 86 View FIGURE 86 ) vs. more than 8.5% TL in other specimens, especially, HUMZ 113578 and HUMZ 117496 (paratypes of S. tokubee ), HUMZ 135083, NSMT 34976, NSMT 61264 (specimens with light spots) and, NSMT 65831, NSMT 65836 and NSMT 65837 (specimens without light spots).
Our analyses indicate that values for mouth width and interdorsal distance in the holotype of S. tokubee (HUMZ 107358) and other paratypes (HUMZ 113574, HUMZ 117472, NSMT 34976) overlap with values of specimens of S. torazame . In relation to mouth width, lower values were found in specimens captured in Tsushima, Sapporo. No differences were found in relation to size of sexual maturity between populations of eastern and western Japan, nor in relation to the color patterns. Additionally, no differences were found in tooth counts, measurements of dermal denticles and neurocranium, and clasper morphology.
Shirai et al. (1992) reported that no specimens of S. torazame were captured along the coast of Shirahama, in the Izu Peninsula. However, according to the specimen catalogue of the Hokkaido University Museum, specimen HUMZ 111033, presenting a color pattern without spots, was captured in Shizuoka Province, the same locality of the types of S. tokubee . This specimen was presented in the list of comparative material of Shirai et al. (1992) identified as S. torazame , with the locality ‘Mito, Ibaraki’ assigned to it. We decided to consider the locality informed by the museum catalogue as it is the oldest record for this specimen.
Shirai et al. (1992: 14–15) pointed out the great variation in color pattern of S. torazame , ‘sometimes showing a petal-like pattern on the lateral surface of the body, but it lacks numerous light spots on the dorsal surface’. Light spots were observed in dorsal surface of many specimens but no quantification was possible in an attempt to distinguish S. tokubee from S. torazame . Therefore, based on the evidence above, S. tokubee is considered here as a junior synonym of S. torazame as no distinctive characters supporting their separation were found.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Scyliorhinus torazame ( Tanaka, 1908 )
| Soares, Karla D. A. & De, Marcelo R. 2019 |
Scyliorhinus tokubee
| Weigmann, S. 2016: 44 |
| Compagno, L. J. V. & Dando, M. & Fowler, S. 2005: 253 |
| Shirai, S. & Hagiwara, S. & Nakaya, K. 1992: 9 |
Scyliorhinus torazame: Fowler, 1941 : 36
| Weigmann, S. 2016: 44 |
| Sinohara, G. & Nakae, M. & Ueda, Y. & Kojima, S. & Matsuura, K. 2014: 232 |
| Compagno, L. J. V. & Dando, M. & Fowler, S. 2005: 253 |
| Randall, J. E. & Lim, K. K. P. 2000: 579 |
| Compagno, L. J. V. 1999: 480 |
| Carpenter, K. E. & Niem, V. H. 1998: 1205 |
| Nakaya, K. & Shirai, S. 1992: 40 |
| Compagno, L. J. V. 1984: 367 |
| Masuda, H. & Amaoka, K. & Araga, C. & Uyeno, Y. & Yoshino, T. 1984: 4 |
| Springer, S. 1979: 144 |
| Nakaya, K. 1975: 15 |
| Ueno, T. 1971: 68 |
| Lindberg, G. U. & Legeza, M. I. 1967: 41 |
| Chu, Y. T. & Wang, Y. H. 1964: 675 |
| Mori, T. 1956: 2 |
| Matsubara, K. 1955: 108 |
| Herre, A. W. C. T. 1953: 12 |
| Mori, T. 1952: 18 |
| Fowler, H. W. 1941: 36 |
Halaelurus torazame
| Schmidt, P. J. 1930: 48 |
| Jordan, D. S. & Tanaka, S. & Snyder, J. O. 1913: 10 |
Halaelurus rudis:
| Jordan, D. S. & Tanaka, S. & Snyder, J. O. 1913: 10 |
| Tanaka, S. 1911: 13 |
Catulus torazame
| Honma, Y. 1952: 139 |
| Kamohara, T. 1950: 4 |
| White, E. G. 1937: 107 |
| Matsubara, K. 1936: 36 |
| Tanaka, S. 1931: 5 |
| Garman, S. 1913: 77 |
| Tanaka, S. 1908: 6 |
Scyliorhinus rudis
| Compagno, L. J. V. 1984: 367 |
| Pietschmann, V. 1908: 699 |
| Regan, C. T. 1908: 457 |