
Xystopyge bentemarieae, Enghoff, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.803.1691 |
|
publication LSID |
lsid:zoobank.org:pub:8B66C8AE-F00A-42F6-9641-26B0ECC49F78 |
|
DOI |
https://doi.org/10.5281/zenodo.6373731 |
|
persistent identifier |
https://treatment.plazi.org/id/0968ADE8-6469-44B3-AE8D-D427EED0A597 |
|
taxon LSID |
lsid:zoobank.org:act:0968ADE8-6469-44B3-AE8D-D427EED0A597 |
|
treatment provided by |
Felipe |
|
scientific name |
Xystopyge bentemarieae |
| status |
sp. nov. |
Xystopyge bentemarieae sp. nov.
urn:lsid:zoobank.org:act:0968ADE8-6469-44B3-AE8D-D427EED0A597
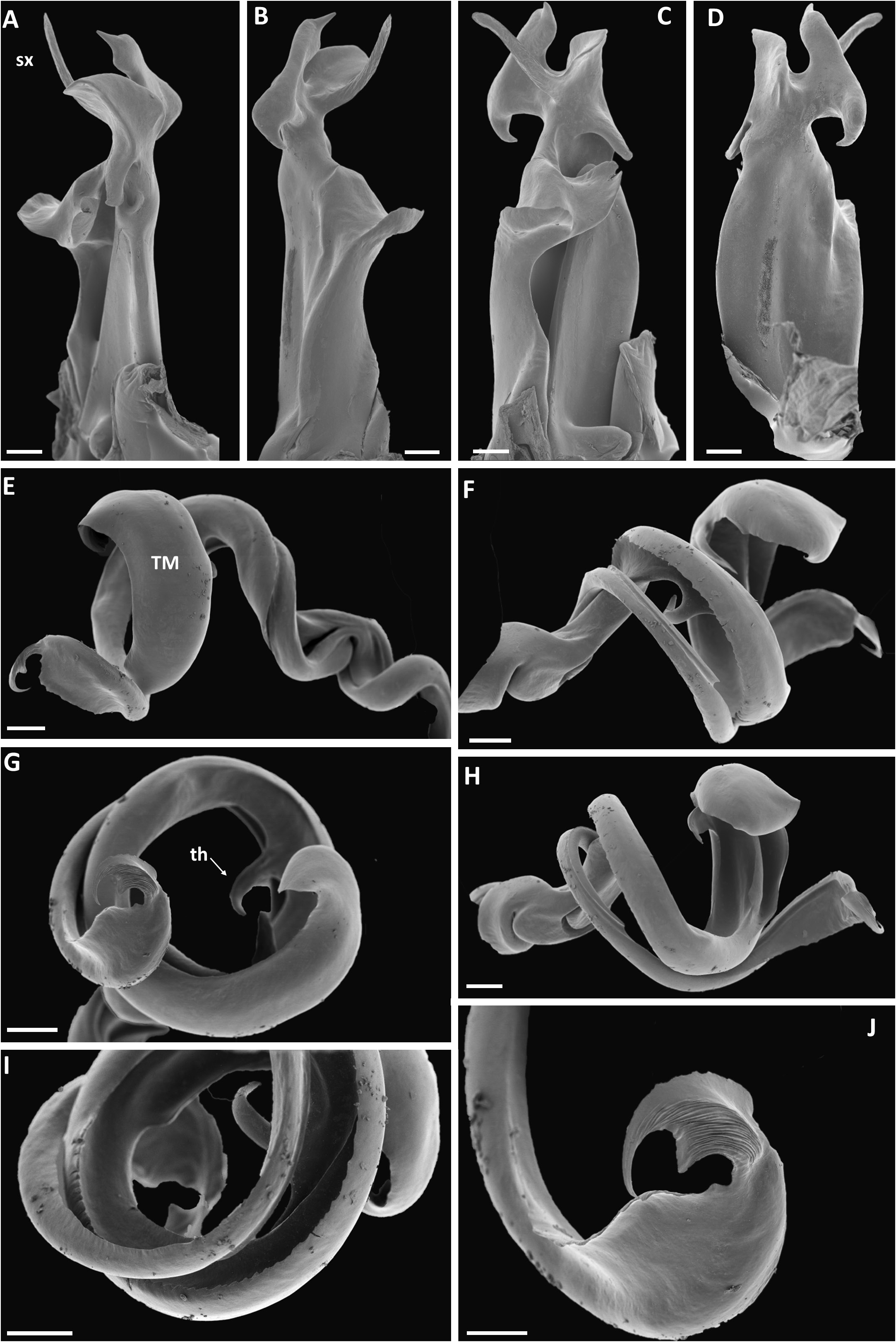
Figs 4B View Fig , 62–64 View Fig View Fig View Fig
Diagnosis
Differs from other species of Xystopyge by the combination of a hammerlike profile of the distal part of the gonopod coxa in certain views ( Fig. 63C View Fig ) (shared only with X. martella VandenSpiegel & Pierrard, 2004 , and X. doggartae sp. nov.), an only moderately long, tongue-shaped gonopod sternum (very long, slender-triangular in X. martella and X. doggartae sp. nov.), a very stout, moderately curved hook (th) at midlength of the gonopod telomere (very strongly curved in X. doggartae sp. nov.), lack of a basal solenomeral spine (BSS) (a small tubercle in X. martella , a long basal spine in in X. doggartae sp. nov.) and a distal division of the solenomere into two branches (undivided in X. martella ).
Etymology
After my granddaughter Bente Marie Enghoff Mogensen.
Material examined (total 7 ♂♂)
Holotype TANZANIA • ♂; Morogoro Region, Kanga Mts , Kanga FR ; 400–500 m a.s.l.; 22–25 Nov. 1984; lowland rain forest; N. Scharff leg.; NHMD 621766 .
Paratypes TANZANIA • 6 ♂♂; same collection data as for holotype; NHMD 621767 .
Description
SIZE. Length 57–73 mm. Diameter 3.5–4.0 mm. 61–62 podous rings; no podous rings in fornt of telson.
COLOUR. After 36 years in alcohol faded to light (greyish) brown; however, a broad mid-dorsal pale stripe still evident.
SUPRALABRAL SETAE. 5–6.
MANDIBULAR STIPES. Distal margin slightly concave, posterior-distal corner almost right-angled.
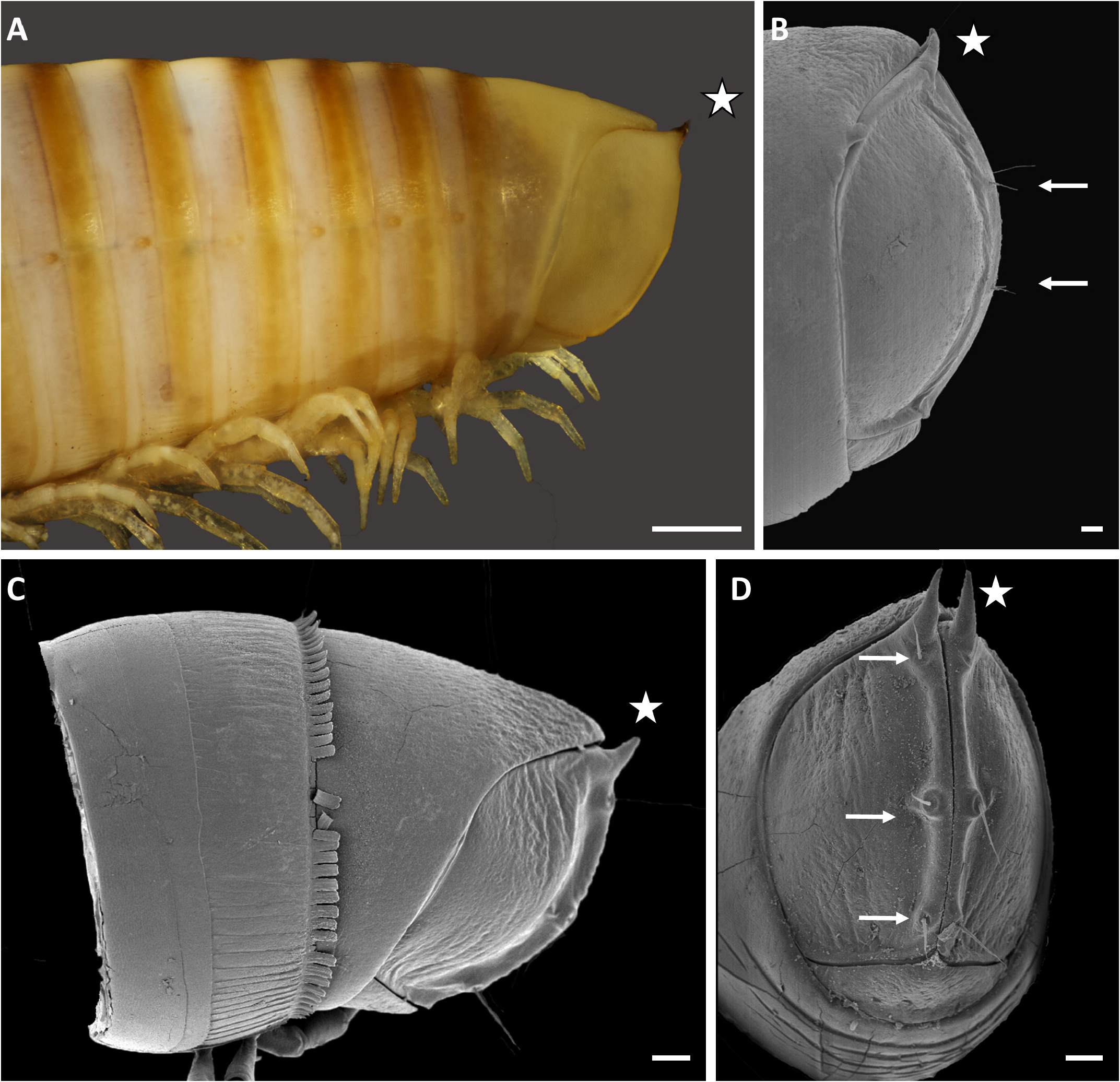
ANAL VALVES ( Fig. 4B View Fig ). Each with a distinct dorsal spine and 2 setae on raised margin; no ventral spine.
LIMBUS ( Fig. 62C View Fig ). Margin with short, smooth rounded lobes.
LEGS. With postfemoral and tibial pads from leg-pair 5, except for last few leg-pairs; size of pads decreasing towards posterior.
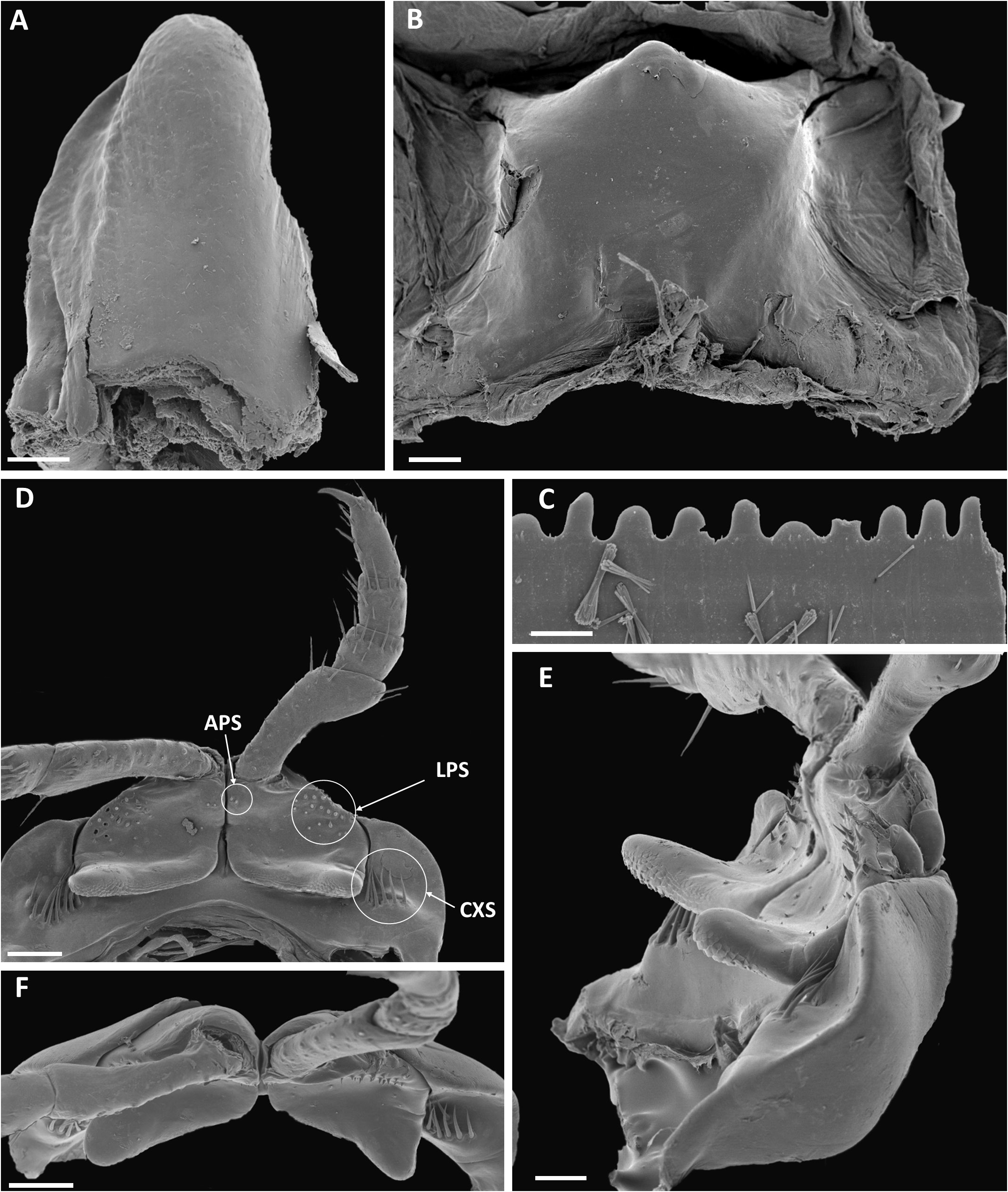
FIRST PAIR OF LEGS ( Fig. 62D–F View Fig ). Prefemoral lobes short, triangular in ventral view. Six to eight long coxosternal setae (CXS) adjacent to lateral side of prefemoral process; prefemur with a few peglike mesapical sensilla (APS) and ca 12 peglike lateral sensilla (LPS).
GONOPOD STERNUM (STERNUM 8) ( Fig. 62A View Fig ). Tongue-shaped, ca twice as long as broad.
STERNUM 9. Pentagonal with parallel lateral margins (‘house-shaped’).
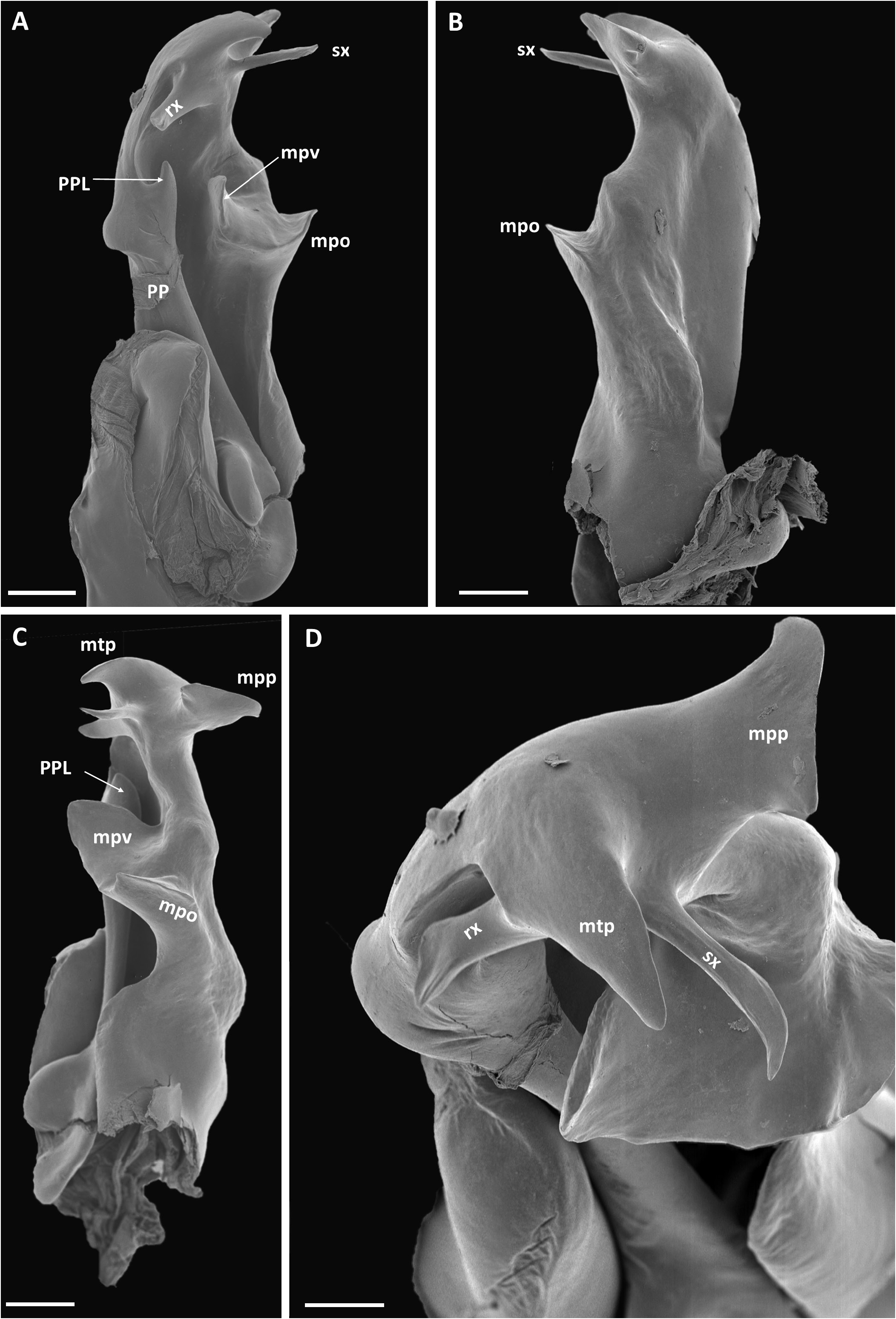
GONOPOD COXA ( Fig. 63 View Fig ). Proplica (PP) simple, proplical lobe (PPL) relatively slender, clearly visible in anterior view. Metaplica (MP) at level of proplical lobe produced mesad into triangular ridged process (mpo), at lateral end of mpo a large vertical lobe (mpv), distally produced into complex structure with four subhorizontal processes: 1. A large, subrectangular meso-posteriad process (mpp), 2. A long, triangular mesad process (mtp), 3. A very slender process (sx) process originating next to mtp 4. A subrectangular anteriad process (rx) (sx and rx corresponding to the similarly labelled processes in X. doggartae sp. nov.).
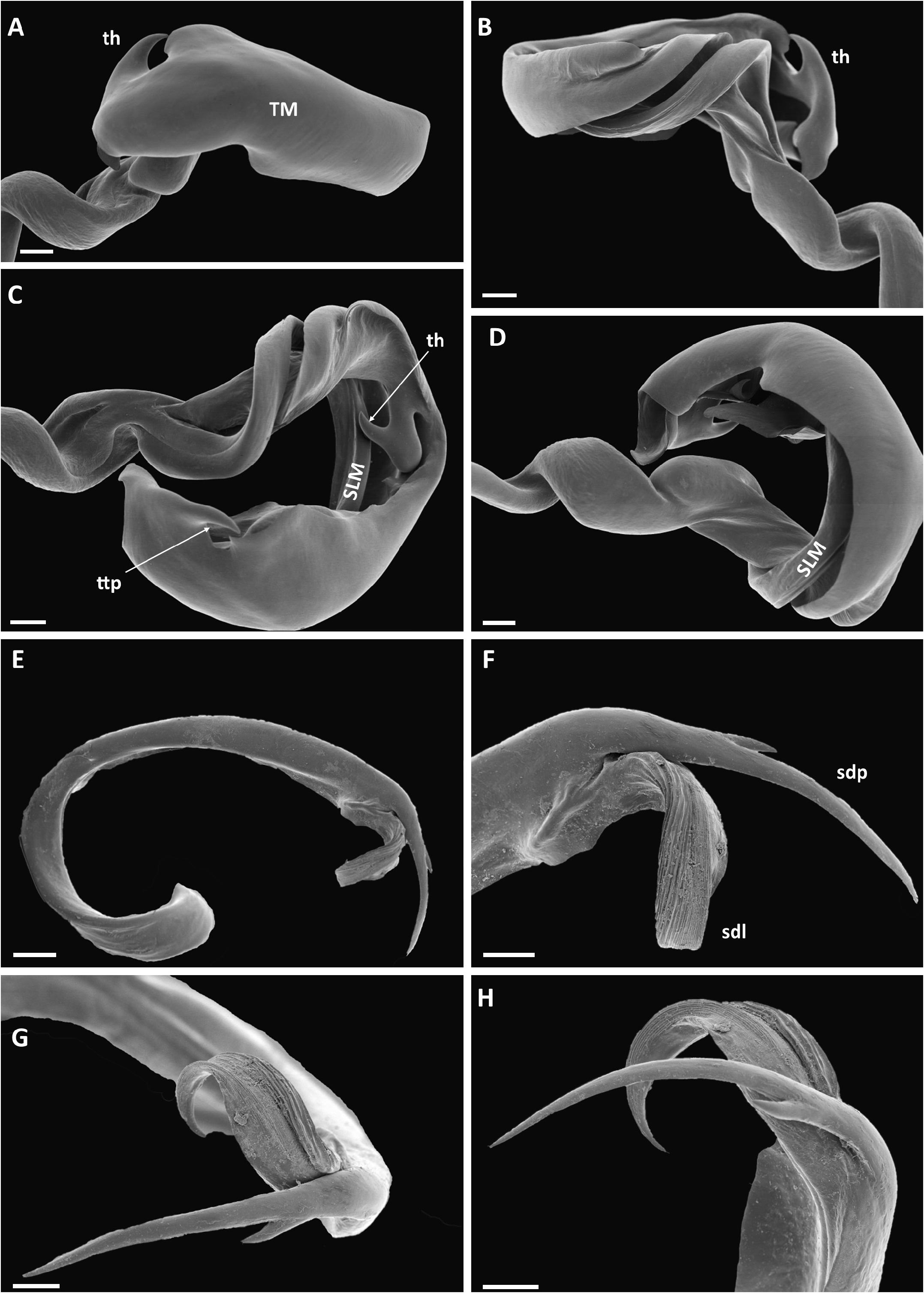
GONOPOD TELOPODITE ( Fig. 64 View Fig ). Arculus 135°. Torsotope not very well delimited. Solenomere (SLM) at rest concealed within gutter formed by telomere; ca as long as telomere, simple, without a basal spine, towards end divided into a long, striate branch (sdl) with the opening of the efferent canal, and a longer, slender, smooth, curved spinelike branch (sdp), the latter with a short basal accessory spinelike branch. Telomere (TM) overall consisting of a ribbon describing a full circle and at the same time folded lengthwise forming a concavity along the inner side of the circle, with a very stout, smooth hook (th) ca at mid-length; at the end with a transverse spinelike process (ttp).
Distribution and habitat
Only known from lowland rain forest at 400–500 m a.s.l. at the type locality, Kanga FR in the Kanga Mts (part of Nguru Mts).
Remarks
Although X. bentemariae sp. nov. is here diagnosed vis-à-vis X. martella and X. doggartae sp. nov., it also shows some striking similarities with X. robusta Attems, 1910 . This species has been illustrated by Attems (1910) and VandenSpiegel & Pierrard (2004), but the opportunity is here taken to show some SEM images ( Fig. 65 View Fig ) of the gonopods of a male of X. robusta collected close to the Uluguru Mts, viz:
TANZANIA • ♂; Morogoro Region, Morogoro, foot of Uluguru Mts , above the university campus; 17 Mar. 1989; hand collecting from ground and from plants; Mahunka and Zicsi leg.; HNHM diplo-03051 (Compared side by side with a syntype of X. robusta (NHMW 9114, Insel Pemba, Chake Chake, Voeltkow leg. and don.) – the gonopods are completely identical.)
Specific similarities between X. robusta and X. bentemarieae sp. nov. include:
- the very complicated distal part of the metaplica, especially the slender process sx
- the simple telomere (TM)
- the hooklike moderately curved telomeral process (th)
Differences include:
- details of the apical metaplical processes other than sx
- the solenomere (slm) which in X. robusta ends in a striate lamella with a terminal hook, vs divided into a striate branch (sdl) and a spinelike process (sdp) in X. bentemarieae sp. nov.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.