
Veromessor andrei (Mayr, 1886)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5206.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CE749F6C-5832-4152-AB4B-6D89ACCDD560 |
|
DOI |
https://doi.org/10.5281/zenodo.7327867 |
|
persistent identifier |
https://treatment.plazi.org/id/039387FD-FFEE-FFDF-7FC5-FE1658EEF839 |
|
treatment provided by |
Plazi |
|
scientific name |
Veromessor andrei |
| status |
|
( Figures 1A View FIGURE 1 , 2–5 6B View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 )
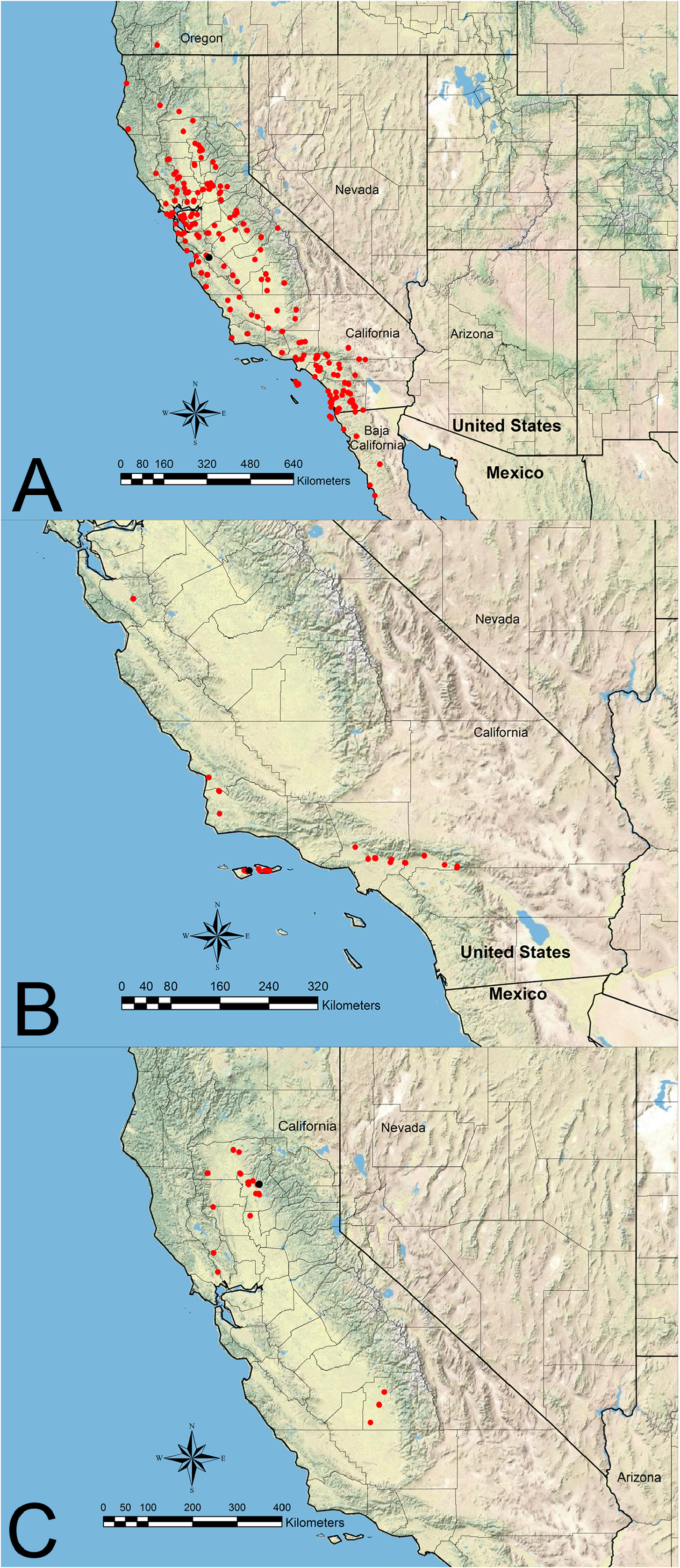
Distribution— Figure 10A View FIGURE 10
Aphaenogaster andrei Mayr, 1886: 448 (worker).
Syntypes examined: 8 workers [NHMW], UNITED STATES, California: no location, April 1884; 1 worker [NHMW], California: San Mateo; 1 worker [NHMW], 12 workers [USNM], California, San Benito County, Tres Pinos; Emery, 1895: 306 (queen); Wheeler and Wheeler, 1972: 240, fig. 7 (larvae); Taber and Colkendolpher, 1988: 95, fig. 1b (karyotype). See also Wheeler and Creighton, 1934: 362, plate II, fig. 2. NHMW worker from Tres Pinos, California, here designated LECTOTYPE [CASENT0923122].
Stenamma (Messor) andrei (Mayr) ; Emery, 1895: 306 (first combination in Stenamma View in CoL [ Messor View in CoL ]).
Novomessor andrei (Mayr) ; Emery, 1915: 73 (first combination in Novomessor ).
Novomessor (Veromessor) andrei (Mayr) ; Forel, 1917: 235 (first combination in Novomessor [ Veromessor ]); Emery, 1921: 67 ( andrei designated as type species of Novomessor [ Veromessor ]).
Veromessor andrei (Mayr) ; Wheeler and Creighton, 1934: 362 (first combination in Veromessor ).
Novomessor (Veromessor) andrei (Mayr) ; Enzmann, 1947: 152 (revived combination in Novomessor [ Veromessor ], in key).
Messor andrei (Mayr) View in CoL ; Bolton, 1982: 341 (revived combination in Messor View in CoL ).
Veromessor andrei (Mayr) ; Ward, Brady, Fisher, and Schultz: 2015: 13 (revived combination in Veromessor ).
Veromessor andrei subsp. flavus Wheeler and Creighton, 1934: 366 (worker).
Syntypes examined: 3 workers [USNM], UNITED STATES, California: San Diego County, Jacumba (W.M. Wheeler leg., 13 August 1917). Creighton, 1950: 159 (synonomy under andrei subsp. castaneus ; here confirmed). USNM worker here designated LECTOTYPE [USNMENT00529212].
Veromessor andrei subsp. castaneus Wheeler and Creighton, 1934: 365 (worker).
Syntypes examined: 5 workers [LACM], 4 workers [USNM], UNITED STATES, California: San Diego County, Jacumba (W.M. Wheeler leg., 13 August 1917). Creighton 1953: 3 (synonomy under andrei ; here confirmed). USNM worker here designated LECTOTYPE [USNMENT00529071].
Worker diagnosis. This species is uniquely characterized by the following combination of features: (1) usually dark reddish-brown to dark brownish, gaster usually darker; some populations in southern California and Baja California light yellowish-orange to ferruginous orange to rust colored; some populations in the Central Valley of California and Monterey area blackish to black, (2) medial lobe of clypeus with 2–3 coarse, lateral longitudinal rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4) entire circumference of base of scape with a strongly developed and flattened, flared and trumpet-like flange; maximum basal width of scape slightly greater than maximum preapical width, (5) MOD less than OMD, OI <23.5, (6) cephalic dorsum with coarse, wavy to irregular, longitudinal rugae that usually have short lateral branches posterior to eyes; medial rugae not diverging to weakly diverging toward posterior corners; cephalic interrugae weakly punctulate, weakly shining, (7) psammophore poorly developed; ventral surface of head capsule with both J-shaped hairs and straight or evenly curved hairs, J-shaped hairs not arranged in a distinct row, (8) in dorsal view, anterior margin of pronotum with one to few irregular transverse rugae, remainder usually with strongly irregular, longitudinal rugae to rugoreticulate; sides of pronotum with wavy to strongly irregular rugae that traverse longitudinally to posterodorsally to rugoreticulate; mesosoma with coarse, strongly irregular longitudinal rugae; mesopleura with mostly longitudinal rugae or rugae angle posterodorsally, rugae (especially on dorsal onehalf) often with lateral branches to rugoreticulate; interrugae on mesosoma weakly punctulate, weakly shining, (9) propodeal spines very slender, acuminate, not curved in profile or in dorsal view; length> 3.0× the distance between their bases, infraspinal facet and propodeal declivity weakly coriarious or with weak irregular, transverse rugae, moderately to strongly shining, and (10) metasternal process large, higher than long, apex very broadly rounded to nearly flat, partly translucent ( Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , 6B View FIGURE 6 ).
Measurements. lectotype (n = 34). HL 1.77 (1.40–1.82); HW 1.70 (1.26–1.81); MOD 0.33 (0.26–0.36); OMD 0.44 (0.28–0.49); SL 1.44 (1.09–1.60); PNW 1.09 (0.84–1.13); HFL 1.89 (1.53–2.29); ML 2.03 (1.82–2.50); PW 0.34 (0.27–0.39); PPW 0.51 (0.40–0.60). Indices: SI 84.71 (79.56–104.69); CI 96.05 (87.07–100.57); OI 19.41 (17.13–23.02); HFI 111.18 (107.75–152.34).
Queen diagnosis. This caste is diagnosed by the following combination of features: (1) usually dark reddishbrown to dark brown with gaster usually darker; some populations in southern California and Baja California light yellowish-orange to ferruginous orange to rust colored; some populations in the Central Valley of California and Monterey area blackish to black, (2) medial lobe of clypeus with 2–3 coarse, lateral longitudinal rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4) entire circumference of base of scape with a strongly developed and flattened, flared and trumpet-like flange; maximum basal width of scape slightly greater than maximum preapical width, (5) MOD slightly less than to slightly greater than OMD, (6) cephalic dorsum with coarse, wavy to irregular, longitudinal rugae; medial rugae not diverging to weakly diverging toward posterior corners; cephalic interrugae weakly to moderately punctulate, weakly shining, (7) psammophore poorly developed, (8) sides of pronotum with irregular longitudinal rugae that often have short lateral branches; mesoscutum with fine, longitudinal rugae, moderately shining; mesoscutellum with longitudinal, oblique, or transverse rugae; anepisternum with longitudinal rugae, katepisternum with longitudinal rugae except for smooth and shining anteroventral margin; interrugae weakly to moderately coriarious, moderately shining, (9) sides of propodeum with longitudinal and oblique rugae; propodeal spines elongate-triangular, length about 1.0× the distance between their bases; infraspinal facet with transverse rugae, propodeal declivity smooth and shining, and (10) metasternal process large, higher than long, apex very broadly rounded to nearly flat, partly translucent ( Figures 7–8 View FIGURE 7 View FIGURE 8 ).
Measurements. (n = 12). HL 1.71–1.89; HW 1.73–1.89; MOD 0.39–0.47; OMD 0.37–0.45; SL 1.41–1.58; HFL 2.03–2.24; ML 2.99–3.38; PW 0.45–0.55; PPW 0.76–0.88. Indices: SI 76.63–88.51; CI 95.60–103.287; OI 22.04–24.87; HFI 115.25–124.28.
Male diagnosis. This caste is diagnosed by the following combination of features: (11) dark brown to blackish, (12) medial lobe of clypeus moderately convex with several irregular, mostly longitudinal rugae, (13) mandibles with 3, rarely 4, small teeth basad of preapical tooth, (14) in frontal view, anterior ocellus above level of top of eyes, (15) mesopleura dull to moderately shining; anepisternum densely punctate, usually with fine longitudinal rugae; katepisternum densely punctate, usually with weak, widely scattered longitudinal or oblique rugae, (16) propodeum rugose to rugoreticulate, interrugae densely punctate; propodeal spines cariniform or with acuminate denticles to small teeth, (17) metasternal process as long as high, tapering to a subangulate to broadly rounded apex, and (18) anterior angle of subpetiolar process subacute to acute ( Figures 1A View FIGURE 1 , 9 View FIGURE 9 ).
Measurements. (n = 12). HL 0.87–1.15; HW 0.79–1.07; MOD 0.34–0.47; OMD 0.10–0.15; SL 0.32–0.43; HFL 1.60–2.27; ML 2.25–2.95; PW 0.30–0.46; PPW 0.53–0.80; AOD 0.11–0.15; IOD 0.28–0.37; OOD 0.30–0.35. Indices: SI 35.42–44.81; CI 84.38–95.54; OI 39.25–47.25; HFI 197.53–233.73.
Additional material examined. MEXICO: Baja California: South Coronado Island, no date (USNM); Coronado Island, no date & Dec 1915 (LACM; USNM); 20 mi N Ensenada, Apr 1, 1931 (USNM); 20 km E Ensenada, 650 m, Apr 27, 1998 (UCDC); 2.5 mi SW Mike’s Sky Ranch, 4700’, May 11, 1998 (RAJC; UCDC); 0.5 mi N El Molino, Bahía San Quintin, Mar 29, 1970 (LACM); Río Amargo (=Arroyo Amargo?), Sep 1, 1961 (LACM); 18 km E Tecate, 850 m, Apr 27, 1998 (UCDC). UNITED STATES: California: Alameda Co.: Livermore, Mar 14, 1937 (LACM; USNM); Hayward, Oct 2, 1910 (USNM); Berkeley, Jun 18, 1917 (LACM); Mills College, no date (LACM); Niles Canyon, Apr 15, 1922 (LACM). Amador Co.: 9 km WNW Plymouth, 200 m, Mar 20, 1994 & Aug 11, 1994 (UCDC); 3 mi SSE Ione, 120 m, Jun 4, 1988 (UCDC). Butte Co.: Bidwell Park, Apr 26, 1975 (LACM); Neal Rd at 6.1 mi NE Jct Hwy 99, 1000’, Jun 23, 1979 (LACM); Pentz, May 1993 (UCDC); North Table Mtn at 9 km N Oroville, 360 m, Apr 18, 1982 & Mar 28, 1993 (UCDC). Colusa Co.: Wilbur Springs, 510 m, Jun 7, 1989 (UCDC); Fout Springs, Jun 27, 1995 (UCDC); 1 km W Fouts Springs, 600 m, May 14, 1983 (UCDC). Contra Costa Co.: Russell Reservation, Aug 29, 1970 (LACM); Mt Diablo State Park, 490 m, Mar 14, 1982 (UCDC); Moraga, Jan 25, 1936 (UCDC); Black Diamond Mines Regional Preserve, 360 m, Apr 10, 1999 (UCDC). El Dorado Co.: 2 km WNW Somerset, 500 m, Aug 16, 1987 (UCDC); Shingle Springs, Nov 1, 1980 (UCDC). Fresno Co.: 12 mi SW Coalinga, Nov 12, 1959 (LACM); Fresno, Aug 1, 1991 (UCDC); Joaquin Ridge at 2 km E Wright Mtn, 1240 m, Apr 17, 1999 (UCDC); 2 km SW Marshall Station, 250 m, May 8, 1999 (UCDC). Humboldt Co.: Bair’s Ranch at Redwood Creek, no date (USNM); Etters Cabin, King Ranch Conservancy Area, 10 m, May 24, 1987 (UCDC). Kern Co.: Rancheria Creek on Piute Mtn Rd (T29S, R33E, Section 23), 4350’, Jun 13, 1971 (LACM); Tehachapi, 3950’, Apr 30, 2018 (RAJC). Kings Co.: Kansas Ave (= Ave 232) at 14.0 mi W Hwy 99, 180’, Oct 2, 2018 (RAJC). Lake Co.: 14 km ENE Lower Lake, 290 m, Jun 18, 1995 (UCDC); McLaughlin Reserve, 19 km ESE Lower Lake, 210–700 m, Oct 29, 2010 & Apr 25, 2015 (MLBC); no loc, Apr 18, 1972 (UCDC). Los Angeles Co.: Claremont, Aug 9, 1895 & Nov 16, 1945 & no date (USNM); 3 mi N Claremont, Nov 28, 1969 (LACM); Santa Catalina Island, no date (LACM; USNM); Avalon, no date (USNM); 1 km WNW Catalina Airport, Catalina Island, 410 m, Apr 15, 2002 (UCDC); Los Angeles, Sept 15, 1886 & no date (USNM); 5 mi S Palmdale, Jun 22, 1931 (USNM); Pasadena, Mar 1, 1919 (USNM); Culver City, no date, 1937 (USNM); Tejon Pass, Aug 19, 1917 (LACM); San Gabriel Mtns at Vasquez Rocks, Jul 17, 1984 (LACM); Westwood Hills, May 12, 1936 (LACM); Pomona, Mar 23, 1927 (LACM); Mt Baldy Rd, 3850’, Aug 14, 2004 (UCDC); I-14 at Red Rover Mine Rd, 3040’, Apr 30, 2018 (RAJC). Madera Co. : Patterson, Apr 3, 1954 (UCDC). Marin Co.: San Rafael, Jul 16, 1922 & Jun 26, 1932 (LACM; USNM); Laguna Lakes, Jan 5, 1929 (USNM); Tamalpais, Nov 12, 1922 (LACM); Copper Mines (=Copper Mine Gulch?), Jan 18, 1958 (LACM). Mariposa Co. : 1.6 mi W Mt Bulliam, 2000’, May 6, 1970 (LACM); 4 km SSE Coulterville, 650 m, Mar 25, 2000 (UCDC). Mendocino Co.: Hopland Field Station, 240 m, Apr 29–30, 1977 & Jul 25, 1982 (UCDC). Merced Co.: Snelling, Jun 18, 1951 (LACM); Great Valley Grassland State Park, Jul 25, 1995 (UCDC). Monterey Co: Hastings Reservation, Jul 2, 1944 (USNM); South Hastings Natural History Reservation, Apr 21, 1941 (USNM); Hastings Natural History Reservation, Apr 21, 1941 (USNM); Monterey, May 31, 1943 (USNM); Monterey at School Hill, May 1, 1943 (USNM); Monterey at Lane-Long Field, June 1, 1943 (USNM); Monterey at Forsman Road, Apr 29, 1943 (USNM); Monterey at King City Road, Apr 27, 1943 (USNM); Salinas, Aug 15, 1988 (UCDC); Mission San Antonio de Padua, 340 m, Jun 6, 1998 (UCDC). Napa Co.: Twin Peaks at 4 km W Aetna Springs, 660 m, May 28, 1988 (UCDC); 5 km ENE Rutherford, 120 m, Apr 23, 1988 (UCDC); 5 mi S Pope Valley, Apr 28, 1973 (UCDC); 5 km NNE Calistoga, Oat Mine Hill Trail, 555–645 m, Nov 26, 2010 (MLBC). Nevada Co.: 7 km SW Grass Valley, 700 m, May 30, 1998 (UCDC). Orange Co.: Modjeska Canyon, Jun 2, 1968 (LACM); Laguna Ranch (= Laguna Beach?), no date (LACM); Santa Ana Canyon , May 13, 1934 (LACM); Pleasants Peak in Santa Ana Mtns, 1140 m, Apr 16, 2002 (UCDC); Limestone Canyon at El Toro Rd, 1.8 mi E Cooks Corner, Jul 11, 1996 (UCDC); 6 km N Lake Wildwood, 200 m, May 31, 2013 (MMPC). Placer Co.: Roseville near PFE & Walerga Rds, 110’, Jul 21, 1992 (RAJC); Roseville at 0.5 mi N Jct I-80 & Douglas Blvd, 180’, Jul 21, 1992 (RAJC); Roseville at Jct I-80 & Douglas Blvd, 170’, Jul 21, 1992 (RAJC). Riverside Co.: San Jacinto, Aug, no year & Nov, no year (LACM); Nightingale, 4000’, Sep 2, 1969 & Apr 18, 1971 (NHMW; RAJC); Reche Canyon at 5 mi S Riverside, 1500’, Jun 1, 1966 (LACM); Joshua Tree National Monument at Queen Valley, 5000’, Jul 1969 (LACM); Joshua Tree National Monument at Lower Covington Flat, no date (LACM); Beaumont, Jan 29, 1956 (LACM); 8 mi N Temecula, Jul 19, 1956 (LACM); 15.5 km NE Temecula, 470 m, May 2, 2016 (MMPC); Lake Skinner, 462 m, Mar 29, 1996 (UCDC). Sacramento Co.: Sacramento, Jul 1931 & Jun 17, 1938 & Jun 18, 1941 (LACM; UCDC; USNM); Antelope Pass, May 19, 1932 (LACM); Orangevale, May 24, 1930 (LACM; UCDC); Rancho Cordova, Apr 16, 1984 (UCDC); Folsom, July 18, 1985 (USNM). San Benito Co.: 5 km S Panoche, 425 m, Aug 17, 1988 (UCDC); Hollister, May 30, 1947 (UCDC). San Bernardino Co.: Bighorn Mtns at Rattlesnake Canyon, 3720’, May 24, 2008 (RAJC); Rialto at Jct Riverside & Sierra Ave, 1950’, May 11, 2018 (RAJC). San Diego Co.: Green Valley Falls, 1300 m, Apr 4, 1993 (CASC); Santa Ysabel Valley , no date (USNM); Santa Ysabel, Jul 21, 1895 (USNM); Carlsbad, no date (USNM); Ramona, no date, 1928 (USNM); Descanso, no date (USNM); East Lake, May 25, 1888 (USNM); 1 mi S Jct La Posta Rd & I-8, 3500’, Jul 15, 2000 (RAJC); 1 mi SE Boulder Oaks, 3300’, Jun 30, 1993 (RAJC); 0.2 mi N Cameron Fire Station, 3300’, Jun 28, 1992 & May 13, 2018 (CIDA; RAJC); I-8 at Buck Springs Rest Stop, 3270’, Jul 17, 1992 (RAJC); San Isidro, 200’, Jul 9, 1952 (LACM); Encinitas, Jul 19, 1956 (LACM; UAIC); Indian Flats, 3800’, Apr 12, 1969 (LACM); Escondido, Jul 14, 1956 (LACM); Point Lorma (=Point Loma?), no date (LACM); Carmel Mountain, 124 m, Nov 5, 1993 (UCDC); Chula Vista (CV2), 160 m, Mar 9, 1996 & May 26, 1996 (UCDC); Chula Vista (Home Depot), 70 m, Oct 27, 1995 & Feb 22, 1996 (UCDC); E end Chula Vista, 160 m, Mar 8, 1996 (UCDC); Chula Vista (Paseo del Rey), 178 m, Oct 23, 1995 & Mar 26, 1996 (UCDC); Chula Vista (Terranova), 78 m, Oct 27, 1995 & Jun 5, 1996 (UCDC); Los Peñasquitos Creek, 25 m, Jul 24–29, 2002 (UCDC); Carney Canyon, 455 m, May 9, 2004 (UCDC); Julian, Jun 19, 1962 & Jul 4, 1962 (UCDC); La Jolla, 113 m, Jul 18, 1963 & May 13, 1987 & Jun 26–Aug 1, 1994 (UCDC); Granite Springs at Rancho Cuyamaca, 4800’, Nov 28, 2003 (UCDC); Kate Sessions Park, 92 m, Dec 16, 1995 (UCDC); Hwy 79 at Oak Grove Fire Station, 2760’, May 12, 2018 (RAJC); no loc, no date & Jun 10, no year (LACM; USNM); Pacific Beach, no date (LACM). San Luis Opisbo Co.: 5 mi NE Santa Margarita, Jun 24, 1963 (LACM); Cholame, Mar 23, 1947 (LACM); Paso Robles, Apr 5, 1988 (UCDC); E of Paso Robles, Jul 11–17, 1999 (UCDC); W side Soda Lake in Carrizo Plains National Monument, 580 m, May 24, 2003 (UCDC); 20 km ESE California Valley, Carrizo Plain Natural Area, 800 m, Apr 19, 1997 (UCDC). San Mateo Co.: Jasper Ridge, 150 m, Apr 27, 2003 (UCDC); San Bruno Mtn, 100 m, Apr 5, 1997 (UCDC). Santa Barbara Co.: N end of Sedgewick Ranch, 610 m, Mar 2, 1996 (UCDC). Santa Clara Co. : Palo Alto, no date (LACM; USNM); Stanford University, Sep 26, 1906 (LACM); Mt Hamilton, May 13, 1914 (LACM); Blue Oak Ranch Reserve, 720 m, Nov 21, 2009 (UCDC); no loc, no date (LACM). Santa Cruz Co. : Santa Cruz, Aug 16, 1968 (UCDC); Castle Rock State Park, 880 m, Apr 18, 2010 (MLBC). Shasta Co.: Enterprise, 175 m, Jul 8, 2001 (UCDC). Solano Co.: Cold Canyon, 120 m, no date & Aug 16, 1981 & Apr 6–Jul 9, 1983 & Aug 19–21, 1984 & Mar 27, 1986 (UCDC); Cold Canyon at 19 km NNW Vacaville, 120 m, Jul 9, 1983 & Sep 30–Oct 21, 1987 & Oct 21–Nov 11, 1997 & Jun 29–Jul 20, 1988 & May 31, 1989 & Sep 25, 1991 & Sep 18, 2010 (MLBC; UCDC); 9 mi W Winters at GL Stebbins Cold Canyon Reserve, May 30, 1992 (UCDC); 11 km W Winters, 120 m, Jun 1, 2013 (MMPC); nr Dozier at 11 mi S Dixon, May 4, 1973 & Apr 4–13, 1975 & Sep 11, 1975 & Apr 8–29, 1977 & May 20, 1977 & Apr 11–27, 1979 & Apr 8, 1983 (UCDC); WL Jepson Prairie Preserve TNC at 13 mi S Dixon, Apr 19, 1971 & Apr 19, 1985 (UCDC); Jepson Prairie at 18 km S Dixon, 10 m, Feb 13, 1983 & Jan 29, 1984 & Apr 4, 1993 (UCDC); Fairfield, Oct 19, 1962 (UCDC); Pleasants Ridge, 530 m, Oct 24, 1982 (UCDC); Gates Canyon road at 9 km WNW Vacaville, 480 m, Mar 22, 2014 (MLBC). Stanislaus Co.: 10 mi SW Turlock, May 12, 1952 (LACM); Del Puerto Canyon at 18 km WSW Patterson, 300 m, May 9, 1997 (UCDC); 20 km WSW Patterson, 310 m, May 4, 2013 (MMPC); Del Puerto Canyon, 300 m, May 29, 1971 & May 7, 2005 (UCDC). Sutter Co.: 7 km NNW Sutter, 360 m, May 14, 1988 (UCDC); 1 km W North Butte, Sutter Buttes, 2000 m, May 28, 2007 (UCDC). Tehama Co.: Proberta, Jul 11, 1962 (LACM); 6 km E Paynes Creek, 1360 m, May 24, 1981 (UCDC). Trinity Co.: Weaverville, Aug 15, 1937 (LACM). Tulare Co.: Sequoia National Park, Jul 17, 1931 (LACM; USNM); Goshen, Jul 6, 1988, (UCDC); Tulare, Apr 26, 1937 (UCDC); Pixley National Wildlife Refuge at 0.2 mi W of entrance, 170’, May 1, 2018 & Oct 2, 2018 (RAJC). Tuolomne Co.: 4 mi N Columbia, Apr 26, 1952 (LACM); 5 mi S Columbia, Apr 29, 1952 (LACM); 3 mi N Chinese Camp, 1300’, Apr 18, 1966 (LACM); 14 km SSW Jamestown, 400 m, Jun 4, 1988 (UCDC). Ventura Co.: Simi Hills, 240 m, Oct 2002 (UCDC); 29 km WNW Stauffer, 1460 m, Apr 20, 1997 (UCDC). Yolo Co.: Hwy 128 W of Winters, May 13, 2001 (UCDC); 6 km SW Winters, 45 m, Apr 13, 1984 & May–Jul 1995 (UCDC); 6 km W Winters, 50 m, Apr 13, 1984 (UCDC); Cold Canyon Reserve at 11 km W Winters, Jun 20–Jul 4, 1990 & Feb 5, 2012 & Apr 8, 2012 (MMPC; UCDC); Putah Canyon, Aug 10, 1970 (UCDC); Brooks, Apr 23, 1986 (UCDC); 13 km W Rumsey, 525 m, Apr 24–May 15, 1993 (UCDC); Grassland Regional Park at 8 km SE Davis, 10 m, May 21, 1997 (UCDC); Elkhorn Ferry, Apr 16, 1952 (UCDC); 6 km WNW Rumsey, 270 m, Apr 15, 2012 (MMPC). Yuba Co.: Sierra Foothill Range at 18 mi NE Marysville, May 7, 1977 & May 1, 1983 (UCDC). No county : no loc, no date & no loc, Jun 1884 (LACM; USNM); Mohave Desert, Jul 1931 (LACM). Oregon: Jackson Co.: Siskiyou Mtns at 6 mi W of Jacksonville, 1500’, Jul 22, 1952 (USNM). Questionable locales (appear to be or are outside of geographical range): UNITED STATES: Missouri: Barton Co.: no loc, May 1920 (LACM). Nevada: Ormsby Co.: no further data (not examined, see Wheeler & Creighton, 1934) ( Figure 10A View FIGURE 10 ).
Etymology. The specific epithet, andrei , honored Jacques Ernest Edmond André, an eminent French hymenopterist.
Discussion. Veromessor andrei is not likely to be confused with any congener. This species is easily diagnosed by its poorly developed psammophore combined with long propodeal spines (> 3x as long as the distance between their bases), mostly monomorphic workers, and coarse, irregular rugae to rugoreticulate on the cephalic dorsum posterior to eyes and dorsum and sides of pronotum. This species is recognized in the field based on colonies that consist of tens of thousands of workers that forage in columns, usually near dusk and dawn, and it usually occurs in grasslands or similar open habitats ( Brown & Gordon, 2000; Creighton, 1953; Wheeler & Creighton, 1934). Veromessor chicoensis and V. stoddardi are the only sympatric species that have a poorly developed psammophore, but both species have short propodeal spines (length less than distance between their bases), strongly polymorphic workers, and cephalic dorsum posterior to eyes and dorsum of pronotum with weak, regular rugae.
Wheeler and Creighton (1934) described two subspecies of V. andrei (flavus and castaneus) from the same locality (Jacumba, California). Veromessor andrei subsp. flavus was described based on its lighter yellow coloration combined with morphological differences that included: (1) narrower head (especially in the posterior one-half) with a nearly flat posterior margin, (2) longer antennal scapes that slightly surpassed the posterior margin as in subspecies castaneus, (3) smaller, narrower petiolar node, and (4) more feeble sculpturing, especially on the mesosoma.
Wheeler and Creighton (1934) also described V. andrei subsp. castaneus largely based on its lighter uniform castaneous brown coloration. Other differences included: (1) less polymorphic with the largest workers only 5.2 mm in length (7.0 mm in typical andrei ), (2) longer antennal scapes that slightly surpassed the posterior margin, (3) tooth and lamella on anteroventral surface of petiolar peduncle reduced or lacking so that the ventral surface appears sinuate, and (4) less deeply impressed propodeal suture.
Creighton (1950) synonomized flavus under castaneus without discussion, except to say that flavus and castaneus cannot both be considered valid subspecies as both occur in the same area of southern California. Creighton (1953) synonomized subsp. castaneus under andrei after examining large series from Cameron Valley, about 20 miles east of Jacumba (type locality). Creighton noted that none of the characters separating these two subspecies (except possibly for color) held up after examining adequate material. He also indicated that differences used to separate these two forms resulted from comparing the smaller workers of castaneus with the larger workers of andrei , and that all morphological differences disappeared when comparing same-sized workers. The only remaining difference was the uniformly light color of castaneus, but Creighton noted considerable intranidal variation in color, such that the light castaneus workers sometimes occurred in the same colony as dark andrei workers. We agree with Creighton (1950) and Creighton (1953) in their synonomizing both subspecies under V. andrei .
As mentioned above, V. andrei displays wide variation in color. Wheeler and Creighton (1934) described V. andrei as “Color variable. In some specimens the entire insect is reddish black, except for the nodes of the petiole which are reddish. In other specimens, only the posterior part of the gaster is black, the first gastric segment castaneous and the remainder of the insect clear, deep red. Not infrequently both the head and gaster are infuscated, leaving only the thorax and nodes of the petiole red.” We also have examined numerous color variants that include concolorous orangish, concolorous dark brownish-red, brown, and black, as well as color combinations that include reddish-orange head and mesosoma with a black gaster, orangish-red head and mesosoma with a orangish-black gaster, and head and gaster black with a reddish mesosoma infused with black ( Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ; see also https://www. antweb.org/ and https://www.asu.edu/clas/sirgtools/veromessor/V%20andrei%20second%20layer.htm).
Color does not correlate with geography except that concolorous orangish workers occur only in southern California and northern Baja California, Mexico. Moreover, several morphological traits are stable in V. andrei , while color varies greatly, such that color cannot be used as a taxonomic character. A molecular phylogeny based on UCEs does not show color morphs clustering and indicates an average amount of genetic variation within V. andrei compared to other species, proving color to be of little taxonomic significance in this species (M.L. Borowiec, unpub. data).
Mayr (1886) described V. andrei listing the type locality as “ California ”. We examined 10 syntype workers, each on a separate pin (NHMW), finding several localities on the labels. The label on eight pins was “ California ”, one label was “Tres Pinos, California ”, and one label was “San Mateo, California ”. Moreover, it appears that Mayr examined syntype workers from several localities, but he chose the type locality as “from California ”, perhaps because all 10 syntype workers were from California. We chose the syntype from Tres Pinos, California, as the lectotype so as to define the type locality. Twelve workers from USNM also had a label that read “Tres Pinos, California, Jun, 1884”. We consider these 12 workers to be part of the Tres Pinos syntype series, and we placed syntype and paralectotype labels on all 12 workers.
Biology. Nests of V. andrei occur in open sites in most soil types and consist of one to multiple entrances with irregularly-shaped entrances up to 5 cm in diameter. Nest entrances usually are surrounded by an irregular gravel mound of excavated soil and chaff that ranges up to 60 cm in diameter (Boulton, Jaffee, & Scow, 2003; Creighton, 1953; Snelling & George, 1979; Wheeler & Creighton, 1934). Workers of V. andrei are mostly monomorphic ( Wheeler & Creighton, 1934).
Colonies of V. andrei have not been censused, but observations indicate that they contain tens of thousands of workers that form foraging columns up to 20 m long, with workers fanning out to forage at the distal end of the column ( Brown & Gordon, 2000; Creighton, 1953; Wheeler & Creighton, 1934); vegetation and debris are removed to construct trails when necessary. Colonies sometimes have more than one foraging column. Colonies forage most days during the foraging season, which typically lasts from late March to late October or early November. Foraging time varies seasonally with changes in temperature: colonies forage during the day when days are cool, they become crepuscular-matinal as temperatures increase, and they forage nocturnally when nights are warm ( Brown & Gordon, 2000; Hobbs, 1985). Foraging columns can revisit the same location for multiple days or they can change direction on successive days such that colonies visit the entire area surrounding their nests ( Brown & Gordon, 2000; Hobbs, 1985). Neighbors also affect foraging direction because encounters with neighbors on one day increasing the probability of revisiting that area the following day; encounters with neighbors resulted in fighting that typically involve relatively few pairs of ants ( Brown & Gordon, 2000). This behavior contrasts with that of V. pergandei , in which foraging columns avoid nearest neighbors ( Ryti & Case, 1988a).
Foraging patterns of V. andrei result from chemicals released by several glands. Secretions from the pygidial gland (primarily n-tridecane) appear to initiate the foraging column ( Hölldobler et al., 2013), while a trail pheromone (primarily 1-phenylethanol) released from the poison gland maintains recruitment to the foraging fan (Plowes, Colella, Johnson, & Hölldobler, 2014). The recruitment effect from the poison gland is enhanced when adding pygidial gland secretions (Plowes, Colella, et al., 2014). Like other large-colony congeners, workers of V. andrei have a large pygidial gland reservoir with a textured tergal cuticle ( Hölldobler et al., 2013).
Mating flights of V. andrei occur in post-dawn hours over an up to several week period from mid-June through July, usually with relatively few sexuals released per day ( Brown, 1999b; McCluskey, 1963; Snelling & George, 1979; R.A. Johnson, pers. obs.). Flights are synchronized across colonies, apparently in response to a circadian rhythym in which activity of males increases drastically during early morning hours ( McCluskey, 1958, 1963). That photoperiod triggers mating flights is based on observations that males exhibit sharp daily activity peaks under controlled light cycles and that these cycles appear to be under endogenous control. Sexuals fly from the nest and mate in the air ( Brown, 1999b; R.A. Johnson, pers. obs.).
Mating frequency for queens of V. andrei is unknown. Dry mass of alate queens averages 7.5 + 0.3 mg. Alate queens contain an average of 44.0 + 1.9 (n = 5) ovarioles, and mated queens contain an average of 1.52 + 0.11 (n = 5) million sperm. Dry mass for virgin males averages 2.1 + 0.1 mg, and they contain an average of 9.15 + 0.64 (n = 3) million sperm (R.A. Johnson, unpub. data).
Little is known about colony founding, but Brown (1999b) observed that founding queens are semi-claustral, i.e., they leave the nest to forage. Brown (1999b) also reported that queens of V. andrei lack storage proteins, inferring that they are obligate foragers, i.e., they cannot rear their first brood of minims without an external food source. Laboratory experiments should examine the founding strategy of V. andrei queens in more detail (e.g., Johnson, 2002, 2006).
Queens of V. andrei also are unusual because dealate queens sometimes occur in foraging columns. Creighton (1953) observed this behavior and suggested that these dealate queens were taken into established nests after mating. These observations were studied by Brown (1999b), who documented that dealate queens occur in foraging columns for about one month, and that all of these dealate queens were uninseminated. The mechanism that causes queens to forego mating and to perform worker-like tasks is unknown, especially in V. andrei , where this behavior cannot be attributed to lack of rains that trigger mating flights. The only other study to examine this behavior in detail found that dealate foraging queens of Pogonomyrmex pima also were uninseminated (Johnson, Holbrook, Strehl, & Gadau, 2007). Dealate queens from mature colonies in several other ant genera also leave the nest to forage (Johnson, 2015; Peeters, 1997).
Most colonies of V. andrei relocate their nest every year, with some colonies relocating nests up to 10 times per year. Relocation was not caused by encounters with their nearest neighbor even though the new nest site was typically more distant from that nearest neighbor. Predation, disease, microclimate, and local resource depletion were suggested as other possible causes of nest relocation ( Brown, 1999a). Patterns of relocation were consistent within this population, but differed from patterns at a distant site ( Pinter-Wollman & Brown, 2015). Veromessor andrei also affects local abundance and distribution of plants through their seed-harvesting and nest building activites. Workers typically display strong preferences for the seeds of some plant species, which results in changing the density and composition of plant species in areas that foragers visit ( Hobbs, 1985; Peters, Chiariello, Mooney, Levin, & Hartley, 2005; but see Brown & Human, 1997). The nest mound itself also consists of a localized microsite that is affected by V. andrei because nest mounds contain higher concentrations of nutrients, a higher abundance and diversity of soil organisms (e.g., fungi, nematodes, microarthropods), and different plant species compared to adjacent non-mound soils ( Boulton et al., 2003; Hobbs, 1985; Peters et al., 2005).
Veromesor andrei occurs in most open habitats from seaside to mountain valleys throughout the California coastal range from southern Oregon to northern Baja California, Mexico, at elevations from 0–2,000 m ( Creighton, 1953; Wheeler & Wheeler, 1973). This species occurs in the Baja California desert, California coastal sage and chaparral, California montane chaparral and woodlands, California interior chaparral and woodlands, California Central Valley grasslands, Northern California coastal forests, and Klamath-Siskiyou forests ecoregions, as defined by Olson et al. (2001). There also are several records from higher elevations (> 1,120 m) in the Mohave Desert ecoregion in San Bernardino and Riverside Counties ( Figure 10A View FIGURE 10 ). We regard the literature record of V. andrei from Ormsby County, Nevada, to be spurious ( Wheeler & Creighton, 1934) until it is reverified.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Veromessor andrei
| Johnson, Robert A., Borowiec, Marek L., Snelling, Roy R. & Cole, Arthur C. 2022 |
Messor andrei (Mayr)
| Bolton, B. 1982: 341 |
Novomessor (Veromessor) andrei (Mayr)
| Enzmann, J. 1947: 152 |
Veromessor andrei (Mayr)
| Wheeler, W. M. & Creighton, W. S. 1934: 362 |
Veromessor andrei subsp. flavus
| Wheeler, W. M. & Creighton, W. S. 1934: 366 |
Veromessor andrei subsp. castaneus
| Wheeler, W. M. & Creighton, W. S. 1934: 365 |
Novomessor (Veromessor) andrei
| Emery, C. 1921: 67 |
| Forel, A. 1917: 235 |
Novomessor andrei (Mayr)
| Emery, C. 1915: 73 |
Stenamma (Messor) andrei (Mayr)
| Emery, C. 1895: 306 |
Aphaenogaster andrei
| Mayr, G. 1886: 448 |