
Urmiatherium polaki Rodler, 1889
|
publication ID |
https://doi.org/ 10.5252/g2011n4a6 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE374D-5767-C96F-FCAA-FA0EFE89F959 |
|
treatment provided by |
Marcus |
|
scientific name |
Urmiatherium polaki Rodler, 1889 |
| status |
|
Urmiatherium polaki Rodler, 1889 ( Figs 20-22 View FIG View FIG View FIG )
HOLOTYPE. — Part of skull ( Rodler 1889: pls 1-4; cast NHML M4114 View Materials ).
TYPE LOCALITY. — Maragheh, Iran (late Miocene).
Kostopoulos D. S. & Bernor R. L.
MATERIAL EXAMINED. — MNHN.F: cranium, MAR3215 ( Mecquenem 1925: figs 12, 13; pl. VIII, fig. 9); partial frontals with the base of horn-core, MAR1359; atlas, axis and cervical vertebrae, MAR3090 ( Mecquenem 1925: pl. VIII, figs 7, 8; pl. IX, fig. 1); left M3, MAR1055 ( Mecquenem 1925: pl. VIII, fig.4; L = 36.2 mm, W = 27.8 mm); right M2, MAR1054 ( Mecquenem 1925: pl. VIII, fig. 3; L = 31.8 mm, W = 25.5 mm); left M1/2, MAR1056 ( Mecquenem 1925: pl. VIII, fig. 5; L = 28.3 mm, W = 25.0 mm); D4-M1 ( Mecquenem 1925: pl. VIII, fig. 6); p2-m3sin, MAR1035 ( Mecquenem 1925: pl. V, fig. 8 as Antilope indet.); p4-m2sin, MAR2968; m2part-m3 dex, MAR2967; p2-m1sin, MAR2976; m3sin, MAR2027. — NHML: a cast (M4114) of the partially preserved holotype cranium described and figured by Rodler (1889: pls I-IV) and stored in Vienna. — HUW: metacarpal MMTT13/2462 ; proximal and distal part of metacarpal MMTT13/1256.
DESCRIPTION AND REMARKS
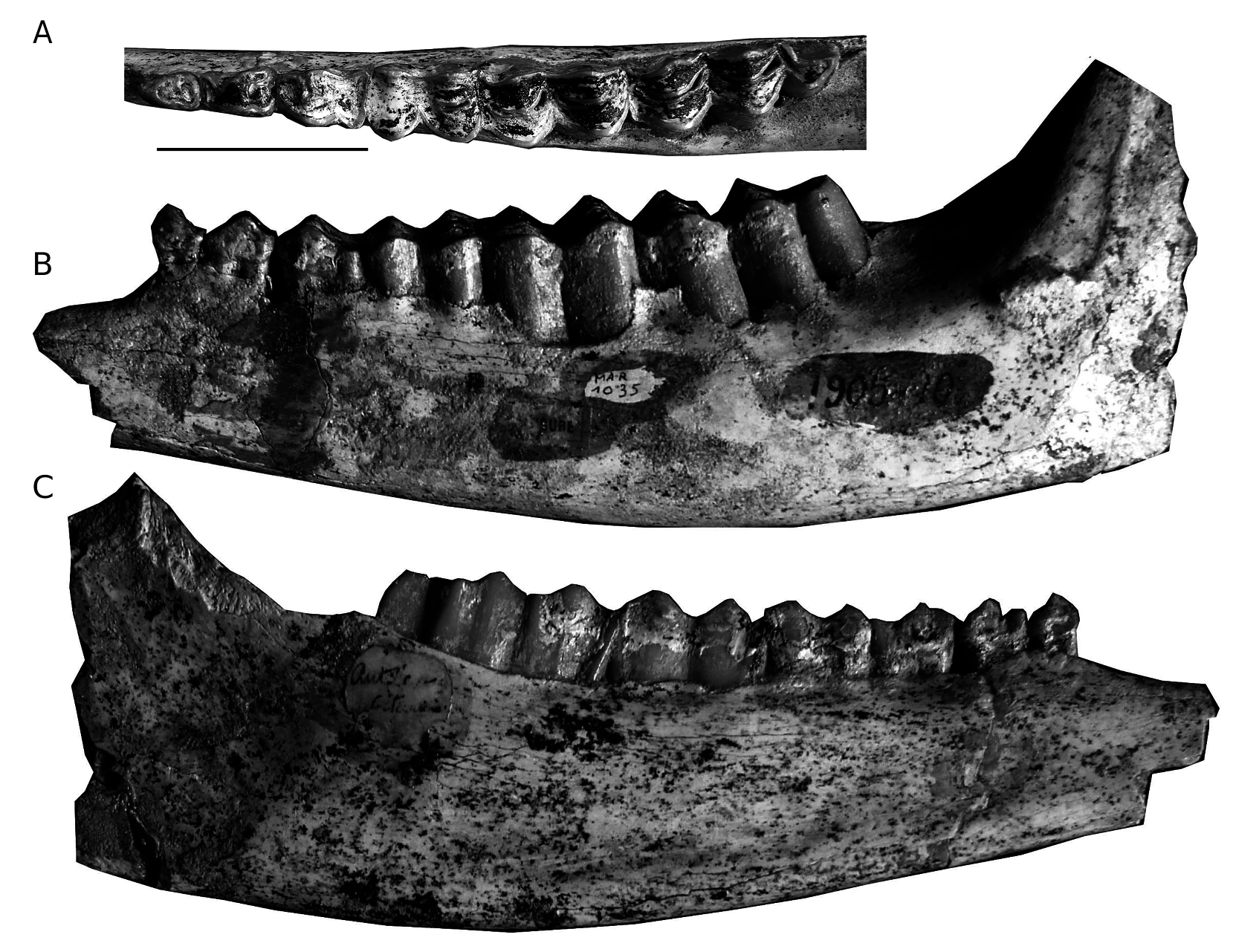
In 1888 A. Rodler announced the discovery of a supposedly sivatherine skull, collected by Dr J. E. Polak from Ilkhchi, SE from Maragheh. In this very brief first note, Rodler (1888) gave some morphological highlights and introduced a new generic name, Urmiatherium . One year later, Rodler (1889) described this cranium in detail as a new species Urmiatherium polaki . Mecquenem (1908, 1925) ascribed to the same species another partially preserved cranium (MAR3215; Fig. 20 View FIG ), several isolated upper molars (MAR1054 -1056) and a few vertebrae (MAR3090; Fig. 20 View FIG ) stored in MNHN.F. Re-examination of the MNHN.F Maragheh collection allows, however, detecting some additional specimens that should be attributed to U. polaki . Urmiatherium is also the most likely referral for two metacarpals in the HUW collection.
The main dimensions of the two available skulls are given in Table 16, though their preservation status does not permit detailed measurements. Mecquenem (1925) suggested that the Paris cranium ( Fig. 20 View FIG ) belongs to an individual ontogenically older than the holotype, possibly because of the shorter and unfused horn-cores of the latter specimen; this is probably correct, although absolute dimensions indicate that NHML M4114 is somewhat larger than MAR3215 ( Table 16). MAR1359 is an additional skull fragment from Maragheh with unfused horn-cores. Since the face is missing from all known specimens, the cranial anatomy of Urmiatherium polaki , already given by Rodler (1889) and Mecquenem (1925), refers only to the braincase and horn-core morphology. Both crania are characterized by stout, short and wide braincase with extremely reduced parietal especially on the top of the cranial roof ( Fig. 20 View FIG ). The enlarged occipital lies on the same level with the parietal and the back of the frontals, altogether facing dorso-caudally. The bulky and round occipital condyles lie on the same plane as the occipital surface and bear large supplementary facets for the atlas articulation. The strong paroccipital processes are placed more antero-ventrally than the condyles and are directed postero-ventrally; their internal faces also have large additional articular facets for the atlas. The mastoid is large and faces laterally. The nuchal crest is very strong and the fronto-parietal suture forms a rough and thick cordon that marks the posterior contact of the horn-cores with the frontals ( Fig. 20 View FIG ). The external occipital crest is short but strong, surrounded by two localized muscular scars with rough limits ( Mecquenem 1925: fig. 12). The temporal fossa is long and deep with salient borders ( Fig. 20 View FIG ). The auditory meatus is round and rather large. The frontals are significantly elevated above the orbits and extensively hollowed with sinuses invading the horn-core base. The interfrontal suture forms a sharp crest in the holotype but it has been covered by the horn-cores in MAR3215. The orbits are rounded. The basioccipital is moderately short, pentagonal in ventral view and very thick ( Mecquenem 1925: pl. VIII, fig. 9); the left half of the basioccipital bone is slightly stronger than the right one in both skulls, indicating a clear asymmetry. The posterior tuberosities project strongly ventrally, forming together an accessory semicircular (or half-oval) facet for the articulation of the atlas. The anterior tuberosities look like elongated crests that surround the basioccipital towards the rear. A deep and narrow furrow runs between the anterior and the posterior tuberosities. The auditory bullae are small and their great axis is oblique to the sagittal plane. The foramen ovale is moderately large and faces laterally.
The base of the horn-cores covers most of the frontals extending from the anterior-most point above the orbits to the fronto-parietal suture ( Fig. 20 View FIG ). The horn-cores are rather short and robust, with a strongly concave posterior face in MAR3215. Deep, wide longitudinal grooves are present on the posterior and lateral horn-core surfaces of MAR3215 and in the medial surface of the holotype. In lateral view, a deep longitudinal furrow ( Mecquenem 1925: 43) distinguishes the anterior from the posterior part of the horn-core, which seem to follow different rates of development. In sub-mature ontogenetic stages ( NHML M4114 View Materials , MAR1359 ), the horn-cores are directed backwards, being weakly curved and having a spindle-shaped cross-section with great basal axes converging to the rear ; at this stage, the anterior part of the horn-cores is probably low, forming an anteriorly extended buttress above the orbit. Further development ( MAR3215 ) allows the horn-cores to expand their bases mostly medially and posteriorly resulting in their fusion along the sagittal plane and the incorporation of the fronto-parietal ridge into the horn-core base ; at the same time, the supraorbital part of the horn-cores increases upwards forming step-like rugged plates that are attached to the posterior part. Bohlin (1935a, b; 1937) further discussed anatomical details of Urmiatherium horn-cores.
All upper molars are characterized by rather thick enamel and an almost square posterior lobe that is more buccally displaced in comparison with the anterior one. The anterior fossette shows a quite strong posterolingual projection. A large central islet is always present. The parastyle is very strong and is directed mesio-buccally. The mesostyle is strong with a deep, concave anterior flange. The metastyle is weaker than the other two styles and is directed distally. The paracone rib is welldeveloped. The cement is present.
The mandibular corpus is shallow ( Fig. 21B, C View FIG ). The lower premolar row is short, representing 47.7% of the molars ( Table 17). The p2 ( Fig. 21A View FIG ) is simple with a weak parastylid, a distally directed metaconid and broad talonid. The paraconid and the parastylid of the p3 are fused into a single anterior stylid ( Fig. 21A View FIG ), even though a parastylid is still present in the less worn toothrow MAR2976. The strongly distally directed metaconid is long, covering the lingual wall of the talonid. The hypoconid is barely recognizable. The entostylid is short. The p4 is very similar to the p3 but with stronger anterostylid (paraconid+ parastylid) and more anteroposteriorly directed metaconid, both differences resulting in a less open anterior valley ( Fig. 21A View FIG ). The posterior part of the lingual wall of the p4 is continuous. The hypoconid is well-developed, protruding buccally. The first two molars bear a weak goat fold, a strong parastylid, a weak entostylid and a gently crenulated
Kostopoulos D. S. & Bernor R. L.
lingual wall ( Fig. 21A, C View FIG ). There is no basal pillar ( Fig. 21B View FIG ). The m3 has a strong parastylid, a well-developed entostylid, and a moderately well developed metastylid that is present in the upper half of the crown attached to the anterior lobe. A goat fold is absent from MAR2967 but it is present in MAR1035 and especially in MAR2027. The third lobe of m3 is buccally displaced, semicircular shaped and with a rather strong posterior stylid ( Fig. 21A, C View FIG ).
Mecquenem (1925: 44-45) gave a basic morphological description of the cervical vertebrae of Urmiatherium , while Sickenberg (1933) and Bosscha-Erdbrink (1978) further discussed the neck anatomy of Urmiatherium . According to them, the tight atlanto-cranial joint including additional facets in the basicranium suggests a low ability for anterior flexion of the skull while the atlas-axis articulation also indicates restricted lateral movements of the skull on the neck, features that are possibly related with ramming intraspecific behavior.
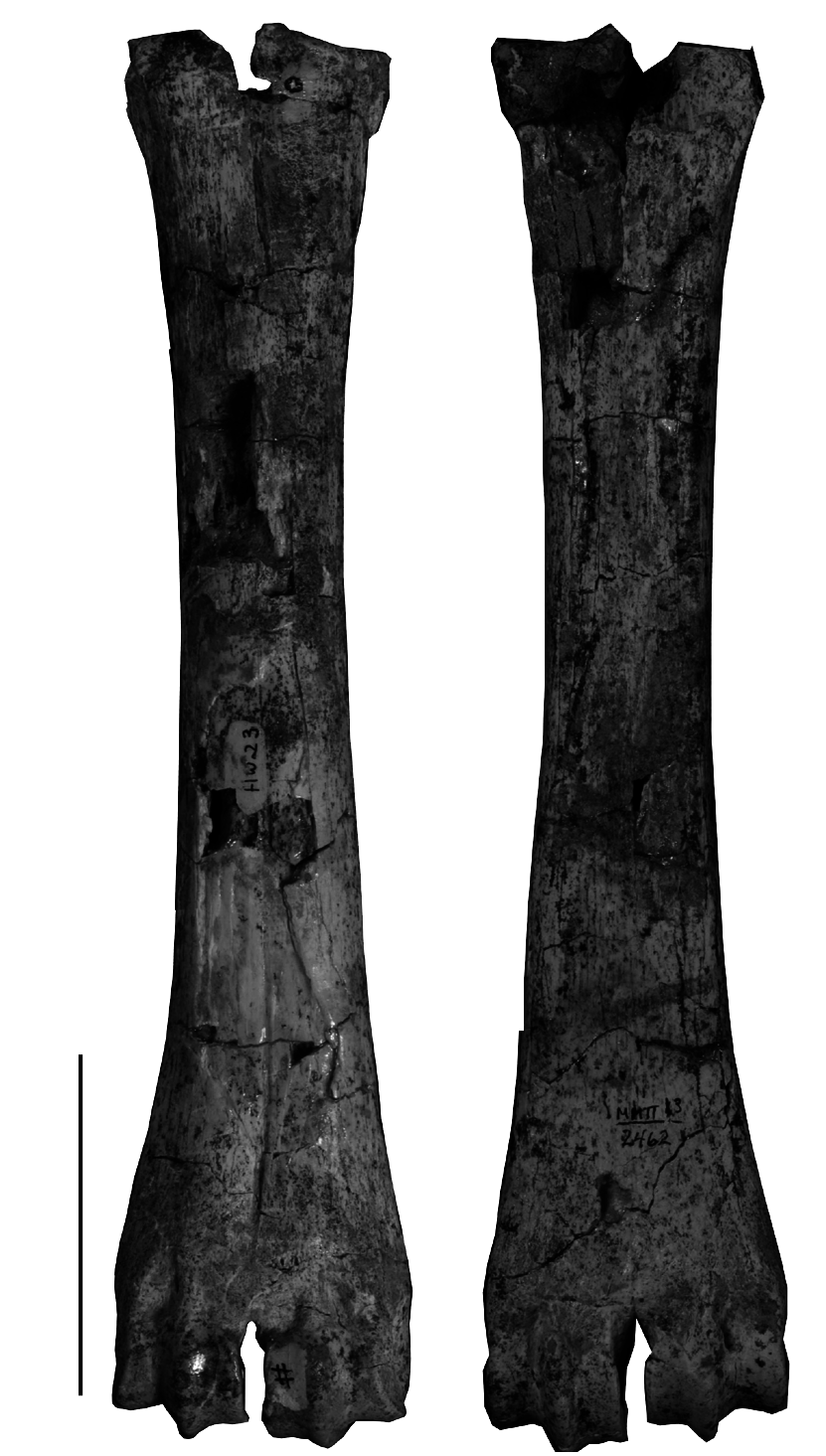
A complete metacarpalIII+IV (MMTT13/2462; Fig. 22 View FIG ) and parts of another metacarpalIII+IV (MMTT13/1256) in the MMTT-HUW collection cannot be associated with any other known bovid species from Maragheh and are consequently regarded as representing Urmiatherium . The metacarpalIII+IV is rather short (210 mm) and moderately robust (the robusticity index DTdiaphysis /Length is 11.6; the index DTdistal /Length is 20.7) with wider distal than proximal epiphysis (DTproximal = 39.5 mm, n = 1; DTdistal = 43.5- 44.5 mm, n = 2). The metacarpal diaphysis has parallel sides and a flat palmar surface. The proximal anteromedial tuberosity is rather weak. The cranial longitudinal groove is reduced. The caudal notch of the proximal epiphysis is open, narrow and shallow. The proximal facet for the capitotrapezoideum is almost squarish. The distal condyles are asymmetric with parallel keels both in the caudal and cranial sides. The supra-articular tubercles of the distal epiphysis are weak and the incisura intertrochlearis is rather narrow.
The Urmiatherium lineage is central to a long discussion concerning tribal affiliations of a set of late Miocene bovid genera referred to as Urmiatherium -group (Gentry 1996). This group exhibits specialized horn-cores and peculiar occipital morphology including – or not – accessory articulations for the atlas (e.g., Bosscha-Erdbrink 1978; Bouvrain & Bonis 1984; Bouvrain 1994; Bouvrain et al. 1995; Gentry 1996; Gentry & Heizmann 1996; Gentry et al. 1999; Geraads et al. 2002; Chen & Zhang 2004; Geraads & Spassov 2008). Apart from Urmiatherium , potential members of this group are Mesembriacerus Bouvrain & Bonis, 1984 from the late Vallesian of Greece, Criotherium Major, 1891 from the early-early middle Turolian of W Anatolia and S Balkans, Plesiaddax Bohlin ,
A B
1935, Tsaidamotherium Bohlin 1935 , and a set of other late Miocene genera from China ( Qiu et al. 2000; Zhang et al. 2000; Zhang 2003; Chen & Zhang 2004). Urmiatherium currently includes three species: U. polaki from Maragheh, U. intermedium Bohlin, 1935 from Shaanxi, China and U. rugosifrons (Sickenberg, 1932) from Samos, Greece (for synonymies see Gentry 1971; Solounias 1981; Bouvrain & Bonis 1984; Bouvrain et al. 1995; Gentry et al. 1999; Kostopoulos 2009a). The Chinese species is known from Locs 30, 43, 44, 49 and 108 of Shaanxi, as well as from Locs
Kostopoulos D. S. & Bernor R. L.
115 and 116 of Kansu suggesting a late Miocene, middle-late Turolian-equivalent age (between 7.2 and 5.5 Ma; Li et al. 1984 and Zhang pers. comm. 2009). Urmiatherium rugosifrons occurs in Samos levels of late middle Turolian age, magnetochronologically dated at 7.1 Ma ( Kostopoulos et al. 2003). “ Parurmiatherium rugosifrons ” from Djebel Hamrin, Irak ( Bouvrain et al. 1995) likely represents a primitive Plesiaddax rather than a small Urmiatherium , while the holotype cranium of Plesiaddax simplex Köhler, 1987 from Kayadibi, Turkey is undiagnostic at the genus level (see also Bouvrain et al. 1995; Geraads & Spassov 2008). Finally, the generic affiliations of Plesiaddax inundatus Bosscha-Erdbrink, 1978 from Garkin, Turkey remain open to discussion (i.e. Bouvrain et al. 1995; Geraads & Spassov 2008).
Since comparison on cranial anatomical details between U. polaki and other related species and genera are already given in Mecquenem (1925), Bohlin (1935a, b, 1937), Bosscha-Erdbrink (1978), Köhler (1987), and Bouvrain et al. (1995) there is no reason to repeat these previous assessments. The new material ascribed to Urmiatherium polaki from Maragheh suggests that this highly specialized large bovid was additionally characterized by a premolar tooth row that is short compared to the molar tooth row and, apparently, moderately elongate and rather stout and wide metapodials.
The upper molars of U. polaki described by Mecquenem (1925) are very similar but slightly larger than those of U. intermedium from China ( Bohlin 1935a), whereas the lower molars of U. intermedium are slightly narrower than those of U. polaki described here. The lower toothrow of U. polaki (Lpm = 123.9 mm) is about 27% larger than that of U. rugosifrons from Samos (Lpm = 98 mm; Kostopoulos 2009a) and about 12% larger than that of Plesiaddax depereti Bohlin, 1935 from China (Lpm = 110-114 mm); it is fully comparable in size with the lower toothrow of U. intermedium from China (Lpm = 102.2- 121.4 mm; Bohlin 1935a) and “ Plesiaddax ” inundatus from Garkin (Lpm = 112.5-133 mm). The reduction of the premolar row seen in U. polaki (47.7% of the molars) is analogous to that of U. intermedium (41.9-49.2%, n = 5, mean = 46.5%; Bohlin 1935a) and Plesiaddax depereti from China (47.6-50.2%, n = 4, mean = 49%; Bohlin 1935a), and significantly more advanced than that of U. rugosifrons (56.1-58.5%, n = 2; Kostopoulos 2009a). “ Plesiaddax ” inundatus shows a greater range but on the average it has longer premolar row (45.3-63.9, n = 11, mean = 53.6; Bosscha- Erdbrink 1978). Criotherium from Samos and Turkey has slightly smaller lower dentition but with longer premolars than U. polaki ( Bouvrain 1994 and pers. obs.).
Morphologically, U. rugosifrons differs from U. polaki in having longer and narrower p2, a p3 with distinct paraconid at first wear stages and more anteriorly oriented anterostylid, shorter and stronger labially protruding protoconid and hypoconid on the lower molars, no goat folds and round, not labially shifted talonid on m3. The mandibular corpus of U. intermedium is rather shallower than that of U. polaki , its p4 is more molarized with contact between the paraconid and the metaconid in 6 out of 8 illustrated specimens ( Bohlin 1935a) and antero-posteriorly oriented metaconid in 5 out of 7 specimens. Its p3 has a subtriangular shaped metaconid, and the paraconid turns backwards in 7 out of 8 illustrated specimens. Plesiaddax depereti has longer diastema than U. intermedium and possibly U. polaki ( Bohlin 1935a) . It also differs from U. polaki in the less tightly fused lobes on the lower molars, the smaller talonid of m3, the much weaker parastylid and the better separated metaconid and entoconid on p4, the forwardly extended metaconid on p3 and the more transverse parastylid of the p3 and p4.
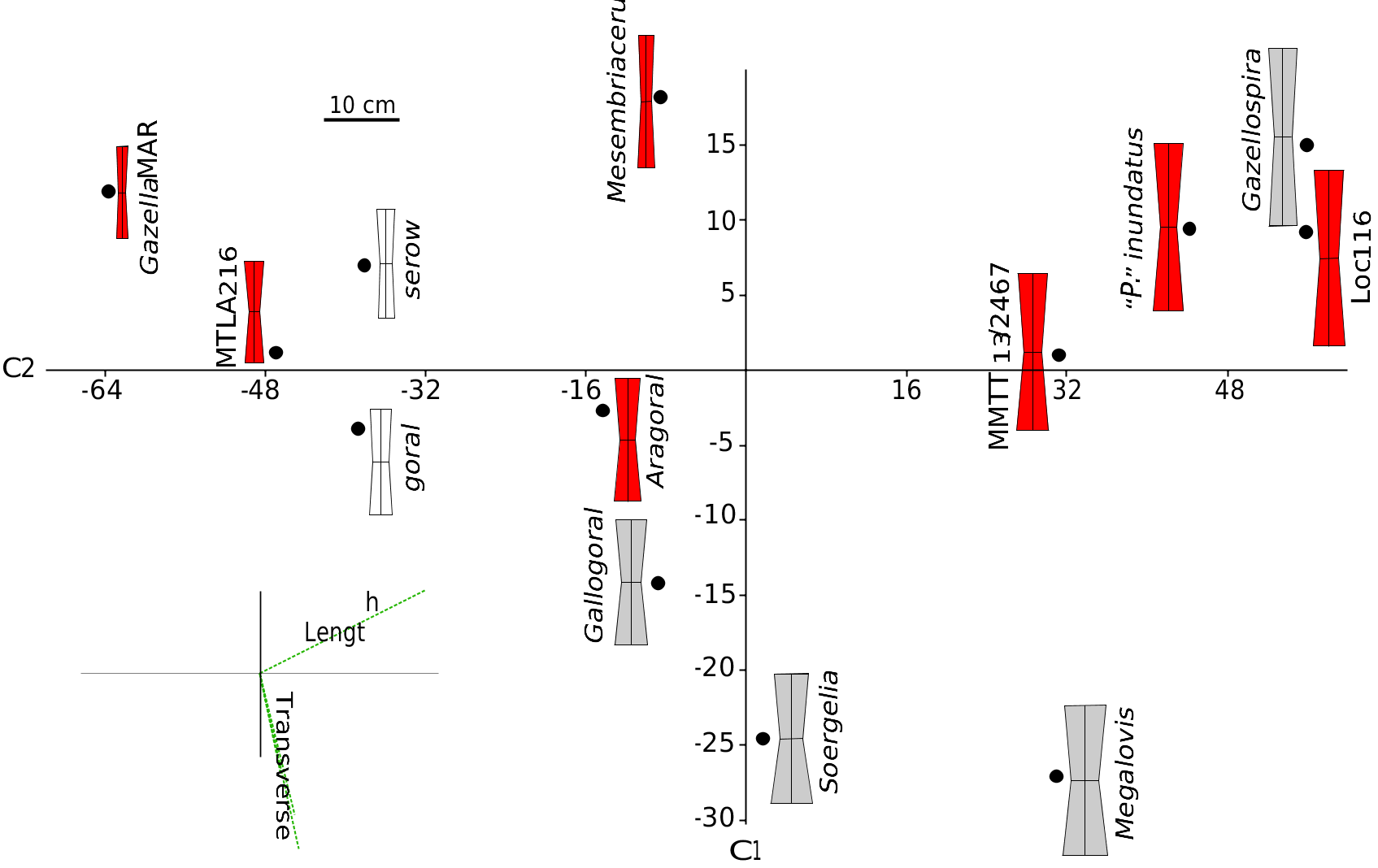
Thusfar, there are few postcranial elements that can be referred to Urmiatherium . Bohlin (1935a) himself refrained from attributing limb bones to the otherwise rich Chinese sample of U. intermedium . Nevertheless, Bohlin (1935a: text-fig. 134) records a rather short canon bone from Baode Loc. 116 that is quite similar to the metacarpals described above and it could be attributed either to U. intermedium or to P. depereti . Recently, Kostopoulos (2009a) ascribed to Urmiatherium rugosifrons some short and wide metacarpals from Samos, which show similarities in their proportions with the metacarpals from Maragheh MMTT 13. Köhler (1993) also illustrated a couple of metacarpals from Garkin, which she referred to “ P. ” inundatus . Comparison of this set of metacarpals with those of several fossil and living antelopes shows that their proportions are intermediate between those of running antelopes and those of heavy Plio-Pleistocene rupicaprines and “ovibovines” ( Fig. 23 View FIG ). The metacarpals of Urmiatherium rugosifrons are close to those of some small sized living Rupicaprini such as serows and gorals ( Fig. 23 View FIG ) probably suggesting similar climbing locomotor adaptations on uneven ground ( Scott 1979). Urmiatherium polaki metacarpals have exactly the same proportions but they are significantly larger. Their length places them close to the Pliocene spriral-horned antelopes but they show broader articular surfaces and shaft suggesting less cursorial adaptations.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Urmiatherium polaki Rodler, 1889
| Kostopoulos, Dimitrios S. & Bernor, Raymond L. 2011 |
Mesembriacerus
| Bouvrain & Bonis 1984 |
Criotherium
| Major 1891 |
Urmiatherium
| Rodler 1889 |
Urmiatherium
| Rodler 1889 |
Urmiatherium
| Rodler 1889 |