
Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890 )
|
publication ID |
https://doi.org/ 10.5252/g2011n4a6 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE374D-5751-C95F-FF4D-FAACFEE4F959 |
|
treatment provided by |
Marcus |
|
scientific name |
Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890 ) |
| status |
|
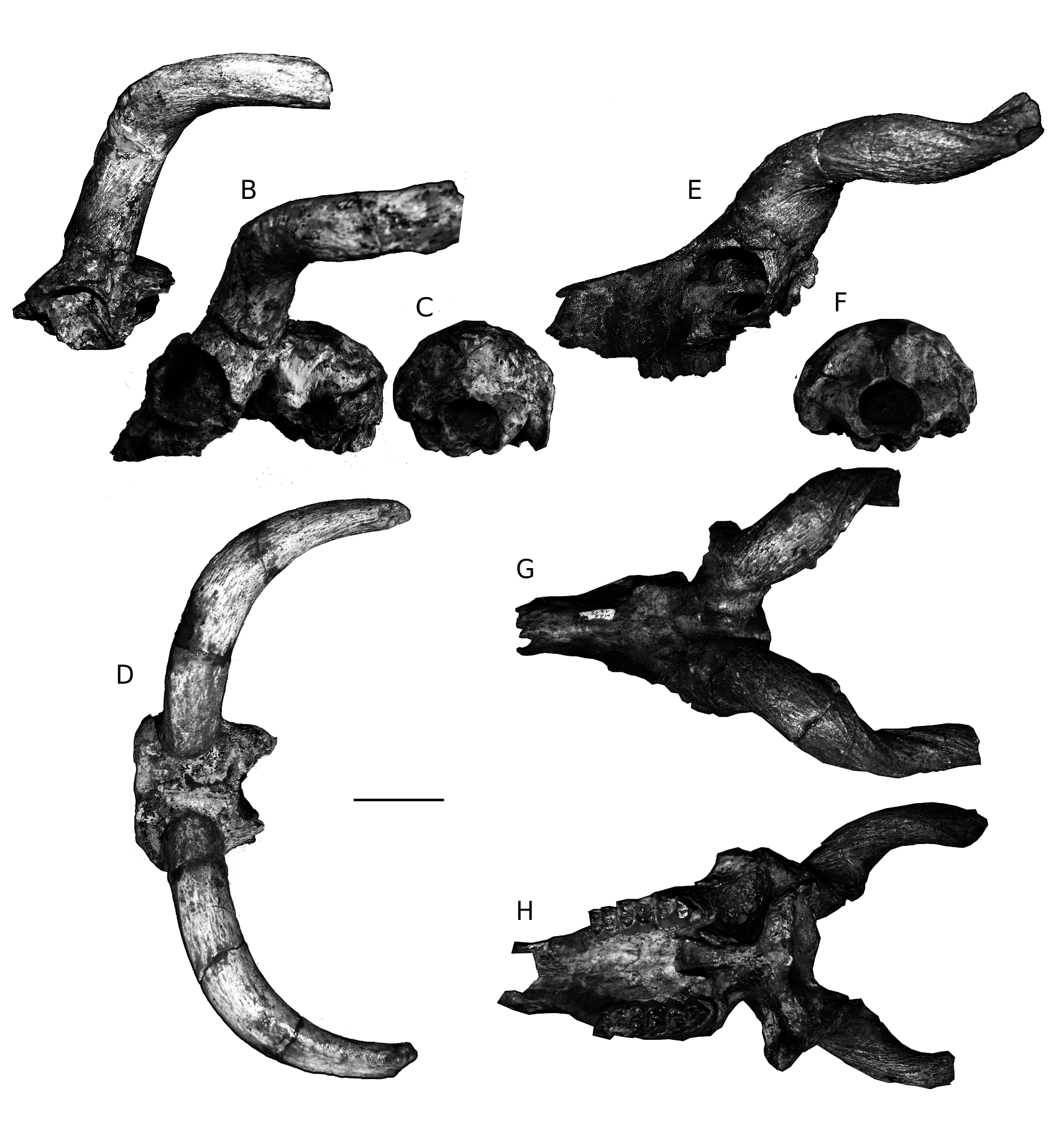
Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890) ( Fig. 13 View FIG E-H)
Tragelaphus View in CoL ? houtum-schindleri Rodler & Weithofer, 1890: 768, pl. 6, fig. 2.
Tragelaphus houtum schindleri – Mecquenem 1925: 37.
Prostrepsiceros houtumschindleri – Gentry 1971: 263 partim.
HOLOTYPE. — Frontlet NHMW 1886-XXVIII.6 ( Rodler & Weithofer 1890: pl. 6, fig. 2).
TYPE LOCALITY. — Maragheh, Iran (late Miocene).
MATERIAL EXAMINED. — MNHN.F: Part of cranium MAR1464 ( Mecquenem 1925: pl. V, fig. 5), 1324, 1312, 1308, 3212 ( Mecquenem 1925: pl. VI, fig. 7); opisthocranium, MAR1061; frontlet, MAR1309, 1831, 1315, 3211, 3213, 3214, 1314; isolated horn-cores, 1819, 1319, 1001, 1816, 1000, 1820, 1818, 1018, 1317, 1815, 1814, 1473; upper toothrows, MAR3173, 3235, 1834; P4-M2, MAR1943, 3237; M3, MAR3238; lower toothrows, MAR1472 ( Mecquenem 1925: pl. V, fig. 7), 1066 ( Mecquenem 1925: pl. VI, fig. 5); p2-m1, MAR3222; p2-m2, MAR3224; p3-m3, MAR3226, 1983, 3216, 1968; p3-m2, MAR3220; p4-m3, MAR3217; p2- p4, MAR3225. — NHMW: holotype, frontlet NHMW 1886XXVIII.6.
DESCRIPTION AND REMARKS
Both Mecquenem’s (1925: 37) and Gentry’s (1971) reports on the Maragheh sample of P. houtumschindleri do not acknowledge its importance as the second most abundant bovid at the site. The species is represented by six crania, seven frontlets, a dozen isolated horn-cores and numerous toothrows ( Tables 4, 5, 9, 10), listing as a whole more than 20 individuals in the MNHN.F collection. Gentry’s (1971) descriptions of this species incorporate characters observed on Samos samples that we do not consider conspecific. Bouvrain & Thomas (1992)
Kostopoulos D. S. & Bernor R. L.
TABLE 9. — Cranial measurements (in mm) of Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890) and Prostrepsiceros fraasi ( Andree, 1926) from Maragheh. For abbreviations see text.
proposed a detailed emended diagnosis of P. houtumschindleri that is largely followed here.
The braincase (MNHN.F. MAR1308, 1324, 1061) is rather wide, short and low ( Fig. 13F View FIG ; Table 9), strongly angled on the face. Its lateral sides run parallel to each other and the dorsal surface is rather flat. The temporal lines are very weak to absent and when visible they converge quickly to the rear. The parieto-temporal suture forms in its posterior end a weak constriction. A deep depression is present just in front of the external auditory meatus, which is small with great diameter trending anteroventrally to posterodorsally. The nuchal crest is well-developed and the occiput semi-oval-shaped. The external occipital crest is blunt and the relevant protuberance is not developed. The occiput faces postero-dorsally and the mastoid mostly laterally ( Fig. 13F View FIG ). The occipital condyles protrude almost vertically from the occipital plane. The basioccipital is narrow and long with strong ridge-like posterior tuberosities and strong, elongated anterior tuberosities trending antero-posteriorly, close together and parallel to each other. A strong crest runs along the mid-basioccipital line between the posterior tuberosities. The foramen ovale is small and faces rather laterally. The auditory bullae are long, not very swollen and their posterior edge fuses with the paroccipital process. The choanae open slightly in front of the lateral indentations of the palate, just at the level of the middle of M3 ( Fig. 13H View FIG ). The orbits are large and round to obliquely ovate with a possible antero-ventral indentation. They do not project significantly on the lateral sides and their anterior rim is placed above the M2-M3 limit ( Fig. 13E View FIG ). The lacrimal fossa is rather shallow without well-defined borders. A thin, long ethmoidal fissure exists. The facial tuberosities are clear and placed above M1-M2 limit. The infraorbital foramina are small, placed slightly above the P3. The premaxillae have a short contact with the nasals, which are long and have weak anterior and posterior lateral flanges ( Fig. 13G View FIG ). The frontals
A between the horn-cores are weakly elevated with a pinched midfrontal suture. The frontoparietal suture is rather complicated and almost perpendicular to the midfrontal one. The supraorbital foramina are sunken into a large and deep depression, the posterior edge of which is placed above the middle of the orbit, i.e. relatively far from the horn-core base (19.5-26.6 mm). The pedicle is very short to absent posteriorly and short anteriorly. A characteristic shallow notch appears anteriorly on the crown of the corneal process.
The horn-cores are inserted on the posterior half of the dorsal orbital rim, being moderately to strongly inclined backwards in their proximal
Kostopoulos D. S. & Bernor R. L.
TABLE 10. — Horn-core measurements (in mm) of Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890) from Maragheh. For abbreviations see text but Hhc is taken here along the anterior surface.
part. They are rather widely spaced at the base; the internal distance between the horn-cores ranges from 22 to 26-rarely to 28 mm. The horn-cores are long, mediolateraly compressed (Table 10), moderately twisted and closely spiralled (1 ½ coils); their maximum uncoiled length probably extended 250-300 mm ( Fig. 13E, G, H View FIG ). The greatest axis of the horn-core base forms a 40-50° angle with the sagittal plane. Seven centimeters above the base, the same axis trends perpendicular to the sagittal plane, whereas 3 cm after, it becomes almost parallel to the same plane. The mean horn-core compression index is 72% at the base (CI = 62- 82%; n = 30) and 135.5% at 7 cm above the base (CI = 125-148%; n = 13) ( Fig. 14 View FIG ). Proximally, the medial surface of the horn-cores is less convex than the lateral one. The posterior keel is always strong, descending postero-laterally above the round to oval and shallow postcornual fossa and associated with a longitudinal furrow along its anterior rim. In most of the studied specimens this furrow disappears towards the horn’s apex. In some of the specimens a set of deep longitudinal furrows descends on the antero-medial edge of the horn-core base, whereas in some others this feature is replaced by a blunt swelling. In several specimens a deep furrow of anteromedial origin is still present in the distal part of the horn-cores. In lateral view the horn-cores curve backwards some centimetres above the base and then run parallel to the horizontal plane ( Fig. 13E, G View FIG ), whereas they re-curve upwards in their distal part. In frontal view the horn-cores diverge moderately in their proximal part and then after they run almost parallel to each other.
The teeth are fairly hypsodont. The premolars represent 56.8-67.2% of the upper and 52.9-58.6% of the lower molar row length ( Fig. 4; Tables 4, 5). The main dental characters of the upper toothrow are the strong paracone rib of the upper premolars and molars that is placed strongly obliquely in P2, P3, M1 and M2, the strong parastyle of P4 and M1-M3, the strong mesostyle and the absence of basal pillars on the upper molars, and the posteriorly protruding metastyle of M3. In the lower dentition, the entoconid of p2 is distinct, the metaconid of the p3 and p4 is elongated and directs strongly distally even though the posterior valley remains as a narrow fissure, the paraconid of the p3 and p4 is strong and vertically placed and their hypoconid protrudes buccally, the lower molars occasionally have basal pillars and they usually have a strong goat fold, and the third lobe of the m3 is singlecuspid and semicircular-shaped.
The homogeneity of the hypodigm’s morphology, as currently understood, as well as the metric dimensions of P. houtumschindleri from Maragheh confirms the species-concept of Bouvrain (1982) and Bouvrain & Thomas (1992) and contradicts earlier observations by Gentry (1971) adopted later by Solounias (1981) and more recently by Bibi (2008), according to which P. zitteli Schlosser, 1904 from Samos is synonymised with P. houtumschindleri . Bernor (1978) referred to P. houtumschindleri several dentitions and horn-cores collected from the Maragheh sites MMTT1A, 1B, 7, 13, 37 and MMTT43 during the LRE. This material is currently housed in Iran and therefore it was not accessible to the first author but the brief descriptions provided by Bernor (1978: 85) allow suggesting that his original horn-core sample includes specimens with quite distinct morphology that probably correspond to different Prostrepsiceros species. Some additional cranial specimens of P. houtumschindleri from Site III of KUE, stored in Kyoto were described later by Watabe (1990: pl. III, figs 3, 4). A morphometrical comparison with relevant Maragheh species is given in the following sections.

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Prostrepsiceros houtumschindleri ( Rodler & Weithofer, 1890 )
| Kostopoulos, Dimitrios S. & Bernor, Raymond L. 2011 |
Prostrepsiceros houtumschindleri
| GENTRY A. W. 1971: 263 |
Tragelaphus houtum schindleri
| MECQUENEM R. DE 1925: 37 |
Tragelaphus
| RODLER A. & WEITHOFER K. A. 1890: 768 |