
Phymaturus manuelae, Scolaro & Ibargüengoytía, 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1939.1.4 |
|
persistent identifier |
https://treatment.plazi.org/id/039C87F6-FFF0-0364-FF37-FE1BFC9AFE85 |
|
treatment provided by |
Felipe |
|
scientific name |
Phymaturus manuelae |
| status |
sp. nov. |
Phymaturus manuelae n. sp.
( Figure 1 View FIGURE 1 )
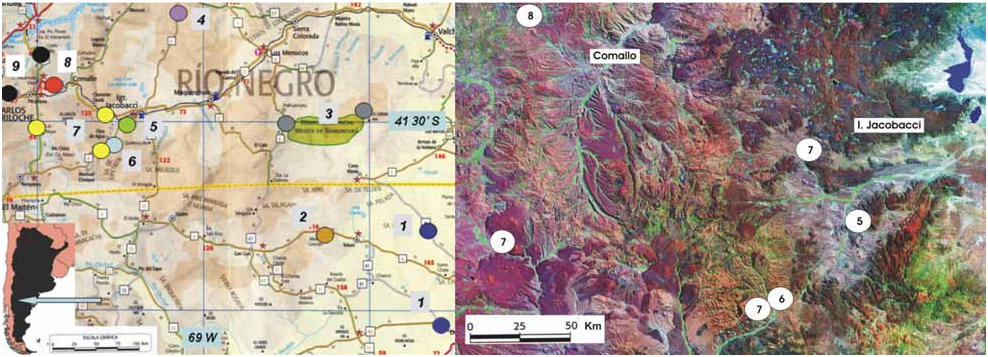
Type Material ― Holotype: MLP-R. 5370, adult male, collected in rocky outcrops (950 m asl) adjacent to National Road 23, 26 km west of Comallo, Rio Negro Province, Argentina. Collected by N. R. Ibargüengoytía, J.A. Scolaro and J. Gutiérrez, 20 December 2007.
Paratypes: UNCo-PH 201, adult male; MLP-R. 5371, adult female; UNCo-PH 202, adult female. The same data as detailed for the holotype.
Etymology ―The species name “ manuelae ” is dedicated to Manuela Martínez the daughter of Nora Ibargüengoytía and Martín, who has accompanied her for most of her fieldwork and congresses since she was born. The name recognizes the help and support of family in our work, so common and valuable to biologists.
Diagnosis ― Phymaturus manuelae shows a noticeable polychromatism. However, as in other species of the genus, females appear to be bigger than males ( Ibargüengoytía 2004). The new species is a member of the patagonicus group, distinguished from the palluma group in having flat imbricate superciliaries rather than being rectangular and non-overlapping; slightly spiny and non-rugose caudal scales in verticilles (as seen among members of the palluma group). However, it has also the subocular scale fragmented (into 2–4 scales; mode = 3) and separated from supralabials by two rows of lorilabials, as in most species of the palluma group, but not seen in the majority of members of the patagonicus group with some geographical exceptions. Phymaturus manuelae may be distinguished from other members of the patagonicus group in having a peculiar coloration.
Description of the holotype ―A medium-sized lizard; snout-vent length (SVL) 90.1 mm; tail 113.7 mm; head length 20.1 mm; head width 17.2 mm; eye-nose distance 5.6 mm; forelimb length measured from to the insertion of the limb into the body wall to the end of the claw of the fourth finger, 33.2 mm; hind limb length measured from to the insertion of the limb into the body wall to the end of the claw of the fourth toe, 52.2 mm; axilla-groin distance 48.9 mm (54.3 % of SVL); fourth finger length 11.8 mm; fourth toe length 14.9 mm; scales in dorsal head 16; scales around midbody 253; ventral scales between mental and precloacal pores 175; scales between rostral and frontal 11; supralabial scales 7-7; infralabial scales 7-7; subdigital lamellae on fourth finger 22; subdigital lamellae on fourth toe 26; precloacal pores 10; cephalic scales subpentagonals, smooth; supraorbital semicircles with large bulky scales, rounded, without azygous, incomplete posteriorly on both sides; no distinct rounded supraoculars; 7–8 imbricate and enlarged upper ciliaries; subocular fragmented in 3-3 almost rectangular scales, shorter than eye diameter, separated from supralabials by 2-2 irregular rows of lorilabials; preocular in contact with first lorilabial row; canthal separated from nasal by one scale; temporals smooth and rounded irregularly pentagonal, in 8–9 scales from auditive opening to the subocular; external auditory meatus enlarged, subtriangular transversally, with 4–5 very protruding or notably enlarged scales on its anterior border; diminute granular scales on posterior border; rostral undivided, wider than higher, separated by one row of small scales from nasals; nasal large and surrounded by seven small scales; nasals separated by three small irregular scales; nostril rounded and large, over the centre of nasal scale; parietals irregular and rough with evident interparietal, surrounded by nine scales; nuchals almost rounded in 4–5 irregular rows; post-auricular folds very developed with round, almost granular scales; mental subpentagonal almost equal in width but higher than rostral, contacted by six irregular rectangular scales; two rows of 4–5 bilateral postmentals decreasing behind; dorsal scales granular, small, rounded and juxtaposed; mid-dorsal scales slightly decreasingly smaller toward ventro-laterals; ventrals larger than dorsals, almost pentagonal, imbricate and smooth; two gular folds with rounded, small scales; 67 gulars between auditory meatus; caudal scales quadrangular and regularly imbricate in verticiles, proximally large and smooth on dorsum, or slightly keeled, distally more rectangular and keeled; scales on limbs round and slightly keeled in the upper side, granular and rounded in the femoral region, larger cuadrangular, imbricate and flat in the anterior and lower side; infracarpals and infratarsals with round margins, becoming keeled to the base of fingers and toes. Subdigital lamellae of fingers keeled; fourth toe and finger claws very developed, almost 2.5 mm of long.
Coloration ― In life, the holotype shows on the head, neck and the greater part of the dorsum (against a black background) an irregular and discontinuous pattern of speckled light-brown spots, more dispersed and attenuated toward the dorso-lateral sides and shoulders (see Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ). On the dorsum from the nape to the base of the tail, appears a group of sub-elliptic spots arranged in two paravertebral discontinuous lines. These spots comprise small, irregular and dense spots of grey or light brown color. The flanks and dorsal surface of limbs are light brown with sparse small and irregular black spots. The dorsal surface of the tail shows a strong variegated coloration featuring ocelli with black borders and light or dark brown centres. Ventrally, chin and throat light gray and speckled by small irregular black spots; these spots are more abundant towards the flanks. The chest is grey and the belly is yellowish or copper-like, more intense toward the cloacal region and the ventral surface of the hindlimbs. On the scales of cloacal region 10 orange reddish colored precloacal pores stand out. The ventral surface of the tail shows attenuate-variegated ocelli.
Morphological variation ―The sample analyzed comprised three adult males and three adult females (see Table 1). Preliminary analyses allow us to establish sex differences: females larger in body size than the males (mean SVL in females = 93.2 mm; range = 81.9–100.5 mm; SD = 9.9; mean SVL males = 89.7 mm). Axilla-groin distance in females ranged 42.5–55.1 mm (mean = 50.8 mm, SD = 7.2) representing 51.9–56.4% of SVL; in males ranged 45.0– 48.9 mm (mean = 46.9 mm) representing 50.3–54.3% of SVL. The total SVL ranged 79.2–100.5 mm. Head length ranged 15.6–20.1 mm representing 19.6–22.7% of SVL. Head width ranged 15.0– 17.6 mm. Eye-nose distance ranged 5.3–6.8 mm. Tail length ranged 94.7–115.5 mm representing 1.15–1.26 times of SVL. Forelimb length ranged 31.1–36.8 mm. Mean of hindlimb length in males was 52.9 mm, but in females ranged 44.3–49.2 mm (mean = 47.3 mm). Scales around midbody ranged 212–253 (mean = 230.5; SD = 16.0). Dorsal head scales ranged 16–18. Ventrals ranged 173–195 (mean = 180.7; SD = 9.0). Precloacal pores only in males ranged 7–10 (mean = 9; SD = 1.5). Fragmented subocular scales in 2–4 (mean = 2.8; SD = 0.6). Two rows of lorilabials between suboculars and supralabials. Scales surrounding interparietal: 6–9 (mean = 7.3; SD = 1.2). Scales contacting mental 4–6 (mean = 4.8; SD = 0.7). Scales between rostral-interparietal: 12–13. Fourth finger subdigital lamellae number: 21–24. Fourth toe subdigital lamellae number: 25–28.
Phymaturus manuelae shows noticeable polychromatism among both males and females, yet all specimens examined possessed the general color pattern described above ( Fig. 1 View FIGURE 1 ). Females show the head, neck and dorsum light brown with irregular black speckled up to the flanks. On the dorsum, eight big and irregular sub-rounded spots are arranged in two light brown paravertebral lines centrally speckled with small dark spots. The tail shows light brown ocelli, with a dark brown spot in the center. Ventrally, the females show the throat, and the anterior zone of the chest grey lightly speckled with small black spots that became bigger toward the flanks. The belly and ventral surface of limbs are grey.
Geographic distribution ― Phymaturus manuelae is only known from its type locality ( Fig. 3 View FIGURE 3 ).
Natural history ―The biotope of Phymaturus manuelae is an isolated basaltic plateau. The peculiar formations are tablelands and flatlands of Tertiary basaltic origin. These tablelands with a predominantly southsoutheast orientation have a very ancient origin, a reduced surface appearing as emergent cones (1000–1200 m asl) isolated in the landscape. They consist of basaltic and repeated depositions of piroclastic material resulting from intense volcanic activity, and are separated by valleys as a consequence of the emergence of the Andes. The deep and narrow valleys are the product of sea ingressions that occurred in the Paleocene with posterior intense hydraulic erosion ( Coira 1979). The physiognomy of those tablelands of similar height shows a plain top with different levels of terraces caused by gravitational slicing and a posterior accumulation of lava and basaltic deposits. The tablelands are separated by few kilometers and their sidewalls of volcanic blocks emerge showing the erosive process that formed these deep valleys. These very fragmented and cracked rocks constitute the biotope of many endemic species of reptiles, and even when they show vicariance phenomena among neighboring tablelands, it is not possible to observe different species at lower altitudes or in the deposits spread in the valleys ( Scolaro 2006).
The biotope is found inside the arid district of Monte Austral, a steppe consisting of open ground, with gravel and effusive rocks. The dominant landscape is the barren steppe, with shrubby, low herbaceous coverage, and the majority of the soil is bare. The dominant vegetation is composed of cushion plants. The dominant floristic physiognomy is low shrubby steppes (with Nassauvia glomerulosa , Chuquiraga oposittifolia , Mulinum spinosum , Senecio filaginoides , Stillingia patagonica , Junellia spp ., Amsinkia calycina , Acaena splendens , Perezia spp ., Ephedra spp ., Fabiana patagonica , Grindelia coronensis , Austrocactus patagonicus ), mean shrubby-grass steppes (with Prosopis denudans , Lycium chilense , Lycium gillesianum , Adesmia patagonica , A. volckmanni , Schinus johnstonii ), and bund grasses ( Stipa papposa , Poa ligularis , Bromus catharticus ) ( Cabrera 1971).
The species inhabits a limited microhabitat that it shares with other Iguanids including some liolaemids ( L. elongatus , L. bibronii , L. ceii ), the leiosaurid Diplolaemus sexcinctus and the gekkonid Homonota darwinii . However, because the new species tends to occupy only the rocky outcrops, few individuals of these other species can be found. The colubrid snakes Philodryas patagoniensis and Philodryas trilineata and the viperid Bothrops ammodytoides are common at the same locality, and together with Diplolaemus sexcinctus may be predators of P. manuelae .
Phymaturus manuelae is viviparous and has been observed during field work eating plants suggesting that, like most members of the genus, is herbivorous.


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |