
Neoscelis dohrni (Westwood)
|
publication ID |
https://doi.org/ 10.1649/604 |
|
persistent identifier |
https://treatment.plazi.org/id/B05C87F4-4672-FFE4-CABF-7C0DFC616F12 |
|
treatment provided by |
Tatiana |
|
scientific name |
Neoscelis dohrni (Westwood) |
| status |
|
( Figs. 1–18 View Figs View Figs )
Larvae. This description is based on 1 cast skin of a third instar larva reared to male pupa, collected in: México: State of Jalisco, Zapopan , Los Guayabos , 29-VI-2001, disturbed oak forest, 5 cm deep in soil with leaf litter and detritus from fungus growing chambers of Atta mexicana, H. E. Fierros-López collector ( IEXA) ; 1 second instar larva reared from eggs laid by 4 females collected in State of Jalisco, San Juan Cosala , 20- IX-1996, G. Nogueira collector ( IEXA) ; 3 first instar larvae reared from the above eggs (IEXA).
Note. Because the cast skin of the third instar is much abraded, and the shape or details of some structures are not clear, the second instar is described and compared with the third and first instars.
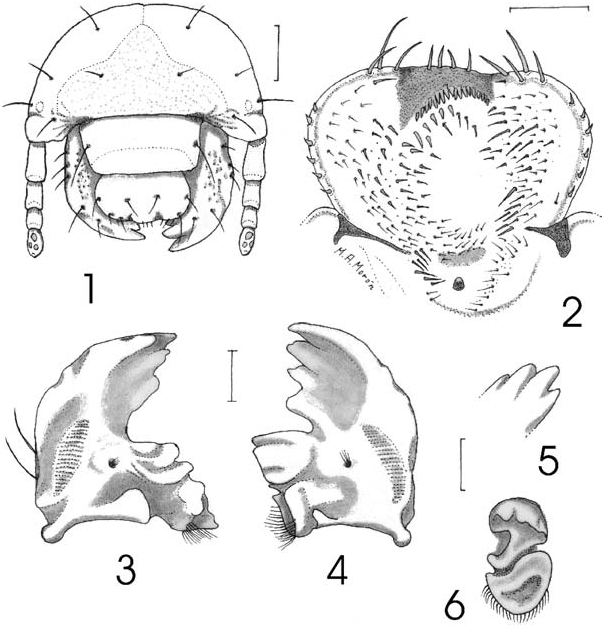
Description. Second Instar Larva. Head ( Fig. 1 View Figs ). Maximum width of head capsule 4.5 mm. Cranium smooth, orange yellowish. Frons with 1 posterior frontal seta, and 1 anterior angle seta on each side, without anterior frontal setae. Dorsoepicranium with 1 dorsoepicranial seta, 1 lateral seta and 1 paraocellar seta on each side. Tentorial pits not defined. Clypeus. Shape subrectangular, with 1 exterior clypeal seta on each side. Preclypeus weakly sclerotized, without setae. Labrum slightly trilobed, clithra present. Epipharynx ( Fig. 2 View Figs ). Corypha with 4 stout, setae. Haptomeral region with wide cone-like process, without macroscopic sensillae, behind process a irregularly curved row of 17 heli, and 4 stout, spine-like setae behind such row. Acanthoparia with 6 short spine-like setae. Chaetoparia with 52–84 setae on each side. Dexiotorma narrowed and long, with well-developed pternotorma. Laeotorma narrowed and short, with well-developed, wide pternotorma. Haptolachus with right nesium weakly sclerotized, with stout setae; sensorial cone small, well sclerotized, with weakly sclerotized area before, and 3–7 setae at each side; crepis vaguely marked. Mandibles. Rigth mandible ( Figs. 3, 5 View Figs ) with 3 scissorial teeth anterior to scissorial notch and 1 scissorial tooth posterior to notch. Stridulatory area elongate-oval, length 3 times its width. Molar area with 3 lobes. Calx wide. Brustia formed by 12 setae. Lateral edge with 2 setae. Left mandible ( Figs. 4, 6 View Figs ) with 2 scissorial teeth anterior to scissorial notch, 1 tooth posterior to notch, and 1 tooth on premolar area. Stridulatory area elongate, length 4 times its width. Molar area with 2 lobes. Acia absent. Brustia formed by 18 setae. Lateral edge with 3 setae. Maxilla. Galea and lacinia fused ( Fig. 7 View Figs ), forming mala. Mala with large uncus at apex and 2 subterminal unci fused at their basis, with a short seta ( Fig. 8 View Figs ); dorsal surface with 7 indistinct rows of setae. Stridulatory area ( Fig. 9 View Figs ) with row of 7–8 curved, acute teeth and a distal, truncate process. Labium ( Fig. 7 View Figs ). Glossa with inverted ‘‘u’’ shaped weakly sclerotized area at middle, with 4 indistinct rows of setae at each side. Hypopharyngeal sclerome asymmetrical, strongly upturned on right side as a tooth-like process with rounded apex; with group of 24 setae on left side; left lateral lobe with 10 setae arranged in 1 indistinct row, right lateral lobe without setae. Antennae. First segment slightly shorter than following 2 segments together. Surface ( Figs. 10–11 View Figs ) of last segment with 3 dorsal and 6 ventral sensory spots. Ocelli weakly defined ( Fig. 1 View Figs ).
Thorax. Thoracic spiracles with C-shaped respiratory plate 0.62 mm high and 0.50 mm wide. Lateral sclerome of pronotum yellowish,wide, with elongate,irregular, central sulcus, surrounded by erect setae.Pronotum with 1 transverse row of 4 long and 2 short setae. Mesonotum with 2 transverse rows of 12–14 mixed short and medium size setae, and 20–24 of mixed medium size and long setae. Metanotum with 2 rows of 20–22 mixed short and medium size setae and 34–36 mixed short,medium size and long setae. Legs. Tarsungulus cylindrical, apex rounded and bearing 9–11 long setae.
Abdomen. Distance between the two lobes of respiratory plates slightly less than the dorsoventral diameter of the bulla. Spiracle of segment I with respiratory plate 0.56 mm high and 0.49 mm wide. Spiracles of segments II–VI similar in size, with respiratory plate 0.60 high and 0.52 mm wide. Spiracle of segment VII slightly larger than preceeding, 0.62 mm high and 0.54 mm wide. Spiracle of segment VIII with respiratory plate 0.60 mm high and 0.54 mm wide. Dorsal areas of segments I–IV each with 3 irregular, transverse rows of 38–62 mixed short, medium size and long setae. Dorsal areas of segments V–VI each with 3 irregular, transverse rows of 17–22 mixed short, medium size and long setae. Dorsal areas of segments VII–VIII each with 23–28 short and long setae. Segments IX and X fused, dorsally with 2 irregular, transverse rows of 29–30 short and long setae. Abdominal segments with long setae 1.5–1.7 mm in length. Spiracular area and pleural lobes of abdominal segments I–VIII with scattered, medium size setae. Raster with pair of palidia each consisting of a irregular row of 17–19 claw-like pali, rows joined posteriorly. Septula narrowed, 8 times longer than wide. Each tegillum composed of 40–44 flattened, apically curved setae. Lower anal lip with mixture of 40–50 long, medium size and short setae.
Third Instar Larva. Similar to second instar except as follows: approximately width of head capsule 6.8 mm; respiratory plate of thoracic spiracles 0.81 mm high and 0.68 mm wide; apex of tarsungulus ( Fig. 12 View Figs ) with 10–11 slender, long setae; respiratory plate of abdominal spiracle of segment I 0.79 mm high and 0.69 mm wide; spiracles of segments II–VI similar in size ( Fig. 13 View Figs ), with respiratory plate 0.82 high and 0.72 mm wide; bulla irregularly oval, slightly convex; spiracles with 38 holes across diameter at middle of respiratory plate; holes irregularly elongated ( Fig. 14 View Figs ); spiracle of segment VII slightly larger than preceeding, 0.85 mm high and 0.74 mm wide; spiracle of segment VIII with respiratory plate 0.81 mm high and 0.72 mm wide; each palidia with 17–20 pali; septula narrowed 6 times longer than wide ( Fig. 15 View Figs ). Thoracic and abdominal chaetotaxy similar, but proportionatelly shorter and sparser than in the second instar.
First Instar Larva. Similar to second instar except as follows: maximum width of head capsule 2.1 mm; epicranial and frontal setae proportionatelly longer; metathorax with minute, spine-like dorso-lateral oviruptor on each side ( Figs. 16–17 View Figs ); tarsungulus cylindrical, 2–3 times shorter than apical or preapical setae; respiratory plates of thoracic and abdominal spiracles kidney-shape ( Fig. 18 View Figs ); respiratory plate of thoracic spiracles 0.16 mm high and 0.11 mm wide; respiratory plates of abdominal spiracles of segment I 0.09 mm high and 0.08 mm wide; respiratory plates of abdominal spiracles of segments II–VIII 0.10 mm high and 0.09 mm wide. Thoracic and abdominal chaetotaxy similar, but proportionately much longer than in the second instar, some abdominal setae with 1.6–1.8 mm in length. Septula and palidia vaguely defined.
Egg. Pearl whitish, ovate, 3.6 mm long, 3.0 mm wide.
Pupa. Male ( Figs. 19–20 View Figs ). This description is based on 1 pupa obtained from the third instar larva collected in: México: State of Jalisco, Zapopan , Los Guayabos , 29-VI- 2001, E. Fierros and J. L. Navarrete ( CZUG) .
Body elongate, robust, exarate. Yellowish white. Surface with very fine velvety microtrichia on last abdominal segments. Head. Strongly reflexed downward; antennae and mouth parts clearly separated; ocular canthus and compound eyes welldifferentiated; labrum tumid; surface of frons and clypeus strongly projected as a widely bifurcate, curved horn-like structure. Thorax. Pronotal disk nearly convex, with irregular shallow depressions toward the sides; lateral margins not defined. Meso- and metanota differentiated. Meso- and metascutellum narrowed backward, with acute apex. Pteroteca widened, free, compressed around body, with two deep, longitudinal sulci, and rounded apex; hind wing teca slightly longer than the elytron teca. Mesometasternal process wide, rounded, emerging between mesocoxae. Protibia without process on external border; meso- and metatibia each with one rounded, short, apical spur; tarsi longer than tibiae, all tarsomeres clearly defined.
Abdomen. All tergites without dioneiform organs; tergite I convex, terguites II–VII with strong, rounded, transverse carinae; tergite VIII nearly convex. Tergo-lateral tubercles II–VI rounded, scarcely prominent. Spiracle I elongate peritreme, not prominent, partially protected by posterodorsal fleshy fold. Spiracles II–IV tuberculiform, with ring-like, sclerotized peritreme. Spiracles V–VII closed, prominent. Spiracle VIII closed, tuberculiform, surrounded by microscopic rugae. Sternites II–VII convex,
10 mm.
with scarce, transverse lines. Last tergite with lateral rounded tubercles, and wide, fleshy lobes on the posterior border, without urogomphi. Genital ampulla wide, trilobed.
Adult Behavior. Most of the following data were obtained from observations, photographs and video recorded in the field by the first author (GN). Males and females show two periods of activity on the host shrubs or trees, between 10:00 to14:00 h, and 17:00 to19:00 h. Males are more active flyers and spend some time flying around the branches at the top of host plants, probably searching for adequate sites for feeding or for perched females that are less conspicuous and apparently more scarce. At the dusk, the males fly from the host plants to the branches at the top of neighboring trees for overnight perching. Males prefer twigs with 1–2 cm in diameter for perching, so that they can embrace the branch completely with their forelegs. Conversely, at the dusk the females fly to the soil under the host plant and bury themselves for the duration of the night.
Both sexes feed on sap flows of Nicotiana glauca , Quercus resinosa , Q. magnoliaefolia , Verbesina greenmanii , Caesalpinia pulcherrima and Acacia pennatula . Three males and one female were found feeding on a single mature fruiting body of Ustilago maydis D.C. ( Ustilaginaceae ). Females seem to spend more time feeding than males and remain for many hours at the same site. Males and females show some preference for special feeding sites because they return daily to the same scars. Apparently the attractiveness of each sap flow increases with the extension of the feeding scar, or with the possible addition of pheromone marks.
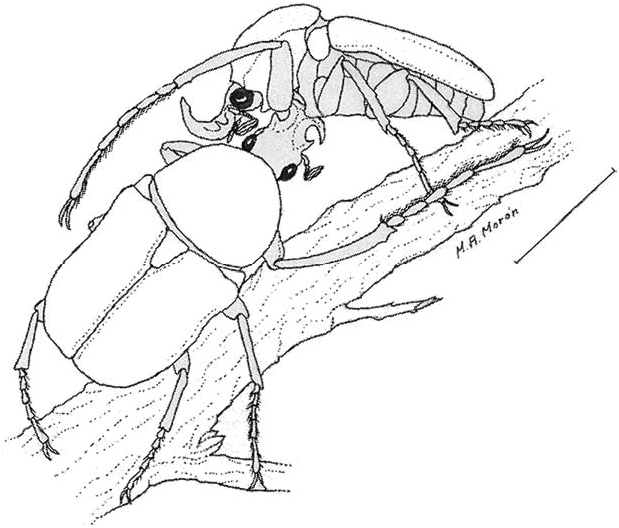
The male walks with rapid movements between the leaves and twigs searching for a female, and when one is found alone, the male quickly mounts the female, surrounding her body and the twig with his long legs. He does not always intend copulation, and frequently the pair have their heads in the opposite directions ( Fig. 21 View Fig ). This behavior probably represents some type of the passive phase of precopulatory guarding (Thornhill and Alcock 1983) when the male is waiting for receptive signals from the female and preventing mounting by other males.
When an ‘‘intruder’’ male approached the precopulatory pair, the ‘‘owner’’ male did not leave the female but did cross some protarsal touching with the ‘‘intruder,’’ and after this the ‘‘owner’’ usually left the female for direct combat with the ‘‘intruder’’.
The first phase of combat behavior involved mainly the very long forelegs of males, that were used to quickly knock over the opponent, without contact of the frontal horns ( Fig. 22 View Fig ). The effect of such knocks apparently is only dissuasive because the dense hair pads on the ventral surface of the tarsal segments do not permit any kind of injury.
At the end of first phase, usually the males briefly make contact with their frontal horns on the head or prothorax of the opponent, and the ‘‘owner’’ quickly returns to the female, covering her body head to head and quickly turning head to pygidium. The first phase takes about 1–2 min, and is repeated over and over until the ‘‘intruder’’ gives up, or both males begin the second phase of combat behavior.
During the second phase of combat, the males make strong and quickly contact with their frontal horns, mainly at the sides or under the prothorax, pushing the opponent off of the twig ( Fig. 23 View Fig ). To avoid being thrown from the twig, the pushed male grasps the pronotum of the opponent with his foretarsi, and remains supported on the opponent without contact with the twig or leaves. The struggle may last 2–3 min or until one male gives up or is pushed from the twig. The ‘‘winner’’ returns to the female and again assumes the precopulatory guarding position awaiting for other ‘‘intruders’’ or for the receptive signals from the female.
During one observation, we saw a ‘‘chain’’ of three males and one female, with the precopulatory pair at the center, one male at the front and the other male at the rear, with the four beetles at the apex of a twig. Aggressive signals between the males were not seen. Other times, the second phase of combat between males was observed without the presence of a female. This may represent defense of feeding sites as cited by Howden and Campbell (1974) and Eberhard (1979) in Dynastinae species that also use the frontal horns and/or enlarged forelegs during combat.
When the female was receptive, the male adopted a head to head position and surrounded the female and the twig with all its legs, holding each tarsus or tibia of the female with his legs before the extrusion of the parameres ( Fig. 24 View Fig ). Copulation took place in this position for 3–5 min. Apparently, the male does not protect the female against other copulatory events afterwords.
Life Cycle. During captivity, females collected at September 20, 1996 died between April 2 and June 2, 1997. Some eggs were laid between October 1996 and March 1997. First instar larvae were observed at May 21, 1997, and second instar larvae were observed at August 24, 1997. Third instar larvae were not obtained from this culture. A full grown third instar larva was collected in the field on June 29, 2001, and a pupa was formed between July 26–27, 2001. Mature adults have been collected in the field between August and October. These data suggest a two year life cycle with overlapped generations.





| CZUG |
Universidad de Guadalajara,Centro de Estudios en Zoologia, Entomologia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |