
Mormopterus eleryi, Reardon, Terry, Adams, Mark, Kenzie, Norm M & Jenkins, Paulina, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.184082 |
|
DOI |
https://doi.org/10.5281/zenodo.5613713 |
|
persistent identifier |
https://treatment.plazi.org/id/03928791-5A0E-FF8F-FDAE-E58EE921FDCB |
|
treatment provided by |
Plazi |
|
scientific name |
Mormopterus eleryi |
| status |
sp. nov. |
Mormopterus eleryi View in CoL sp. nov. Reardon & McKenzie ( Figures 4–10 View FIGURE 4. M View FIGURE 5 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
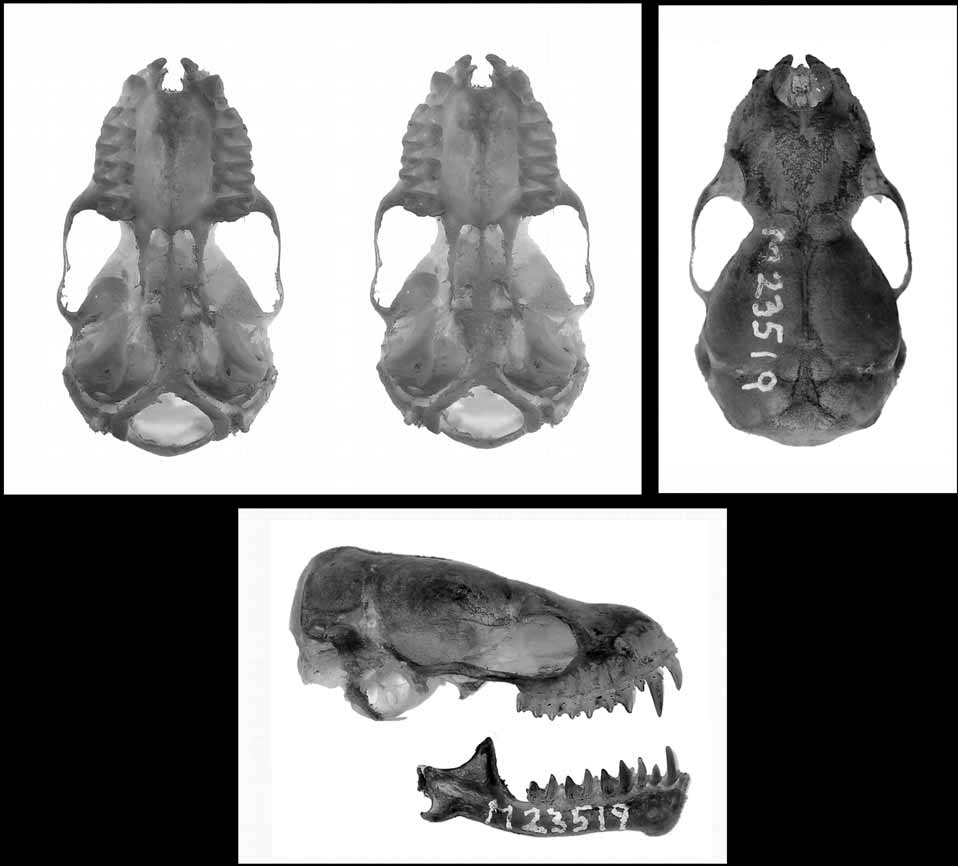
Bristle-faced free-tailed bat (Previous common name: Species 6 ( Adams et al. 1988); hairy-rostrum freetail bat) Holotype. South Australian Museum M23519 View Materials , adult female; body preserved in 10% formalin and then in 70% ethanol, skull extracted. Liver tissue (number ABTC87979) is held in the Australian Biological Tissue Collection at the South Australian Museum.
Type locality. 1.1 km ESE of Eringa, South Australia 26.29184 0 S 134.739030 0E, approximate elevation 240m. Captured by a Constantine trap on the 8th May 2005. The trap was set in a stream flood-out, in Acacia cambagei low woodland over Eriachne ovata , Aristida contorta with emergent Eucalyptus coolabah . Collectors: Jeff Foulkes, Robert Brandle and Lynette Queale (Department of Environment and Heritage, South Australia).
Paratypes. South Australia: SAM M23515 View Materials φ spirit with skull in situ, (Eringa 26.28474 0 S 134.72471 0E); SAM M15811d spirit with skull extracted, Boomika Dam (26.0583 0 S 135.5083 0E).
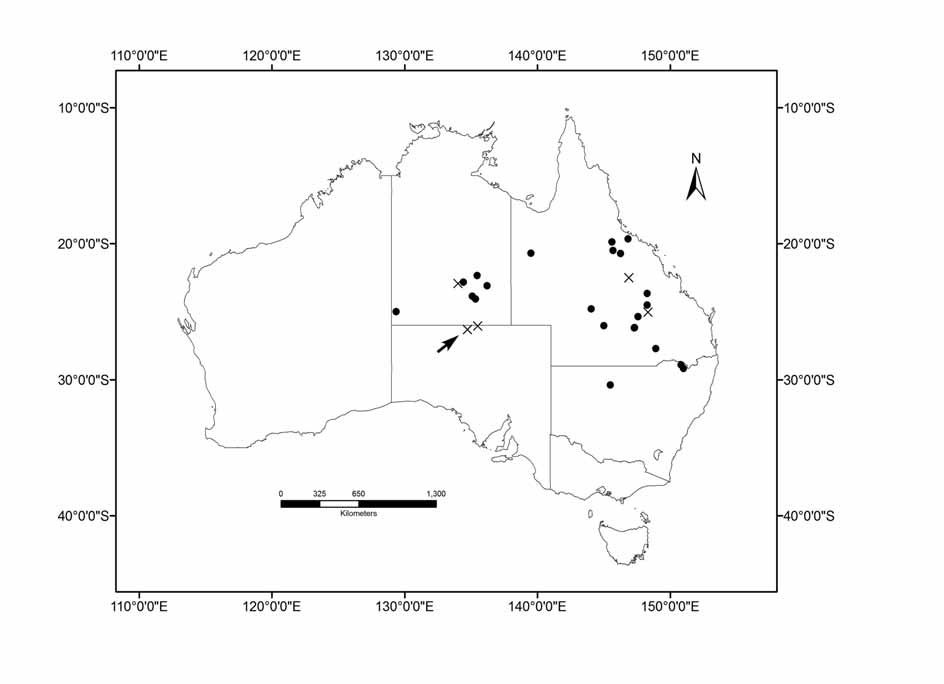
Referred specimens. Northern Territory: WAM M50094 View Materials φ (Bushy Park 21.266 0 S 139.716 0E); ANWC M02427 View Materials d (Alcoota 22.83333 0 S 134.45 0E); NTNM U1043φ (Ringwood Station 23.88 0 S 135.1 0E); U1060d (Dulcie Ranges 22.35 0 S 135.483 0E); U1706d (Ingawallumum 24.08 0 S 135.35 0E); U2734d (Petermann Ranges 25.02 0 S 129.383 0E); AM M9812? (Plenty Downs 23.10 0 S 136.233 0E).
Queensland: ANWC M10448d (Woodstock 19.66 0 S 146.85 0E); QM JM2521d JM2522φ (Fletchervale Station 19.25 0 S 145.817 0E); JM2523d (Helenslee Station 20.517 0 S 145.7 0E); JM2815φ (Warrigal 23.66 0 S 148.28 0E); JM 10367d (King Tank 26.05 0 S 145.016 0E); J20373 View Materials d, J20374 View Materials φ (Mt Owen Station 25.366 0 S 147.6 0E); JM12510d (Idalia National Park 24.8 0 S 144.666 0E); JM11216φ (Nosnillor Station 20.733 0 S 146.266 0E); EBU Q27φ, EBU Q28φ (Beresford Station 22.507 0 S 146.906 0E); EBU Q30d (Early Storms Station 25.0501 0 S 148.3448 0E).
New South Wales: AM M36364 View Materials φ(Bebo State Forest 28.925 0 S 150.835 0E); M33635d (Kwiombal National Park 29.18 0 S 151.01 0E); M33936d (Gundabooka National Park 30.4 0 S 145.5 0E); M50908 View Materials d (Bonshaw 29.05 0 S 151.28 0E).
Etymology. Named after Elery Hamilton-Smith in recognition of his contribution to bat research and conservation.
Diagnosis. Allozyme profile. M. eleryi differs at an average of 49 %FD from M. norfolkensis and at an average of 45 %FD from all other known Australian Mormopterus based on 40 allozyme loci ( Table 2b View TABLE 2 b ). M. eleryi is characterized by alleles not found in any other Australian Mormopterus species at the following nine loci, Ada, Adh 1, Ca, Enol, Got2, Gpd, Gpi, Ldh2, Mpi and PepA ( Table 2 View TABLE 2 b a).
FIGURE 6. Stereo pairs of the holotype of M. eleryi . Top: oblique view of the upper tooth rows. Middle: detail of M1 (left) and M2 (right). Bottom: stereo pair of lower left tooth row.
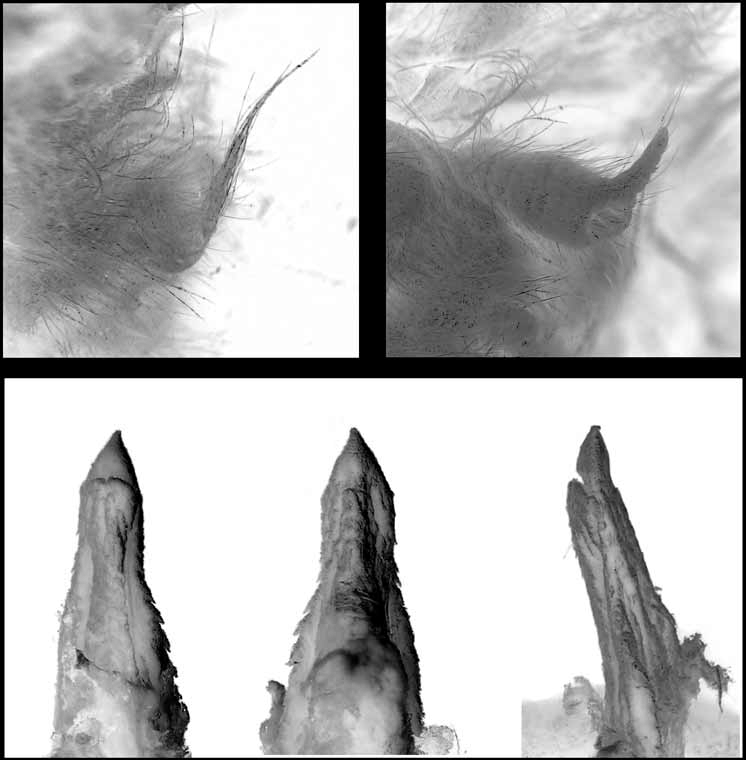
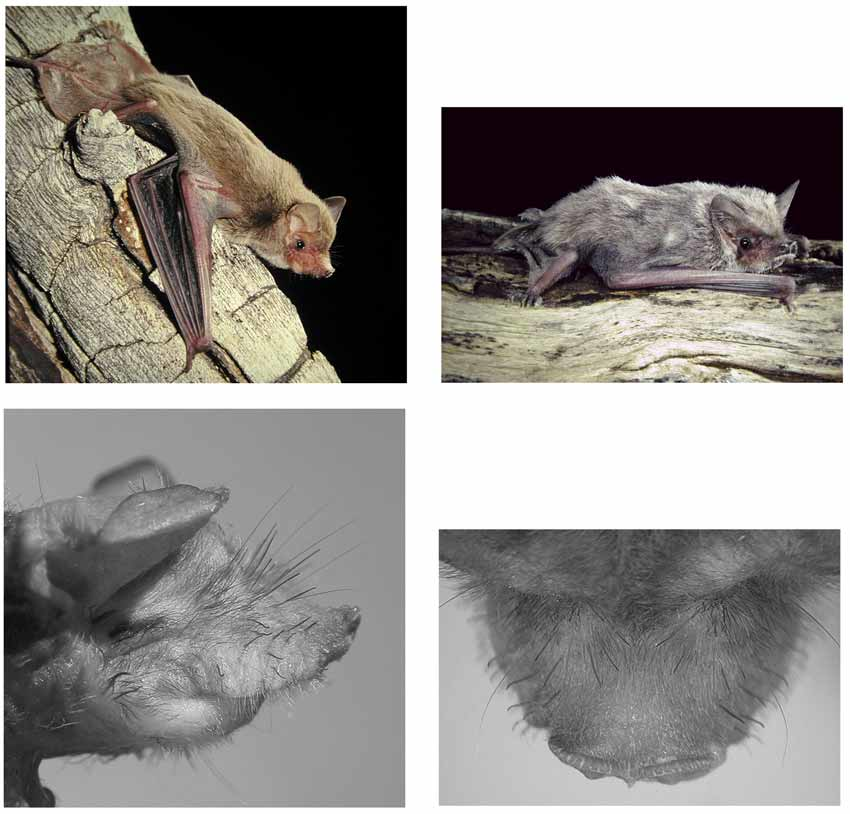
Morphology. M. eleryi differs from all other Australian Mormopterus by the combination of its small size and possession of series of conspicuous stiff bristles arising from the side of the muzzle and face ( Figure 4 View FIGURE 4. M ). The bristles, numbering 26–30 on each side, are shortest (0.5mm) nearer the lips and are longest towards the top of the muzzle (~ 1.6mm). The shorter lateral bristles are sometimes hooked and flattened or spatulate at the tips while the longer bristles are straight and either pointed or slightly flattened at the tips. Bristles also occur in M. norfolkensis but they are uniformly short and less conspicuous.
M. eleryi differs from all other Australian Mormopterus species except M. norfolkensis by the possession of long slender fleshy genital projections in both sexes ( Figure 5 View FIGURE 5 ). The male projection arises from the distodorsal surface of the prepucial skin; the projection length (1.19–1.93mm) averages 43% of the length of the penis. In M. norfolkensis the length of the male projection is 1.1–1.98mm while in the “ planiceps-beccariiloriae complex” it is absent or never exceeds 0.8mm.
The female projection in M. eleryi arises from the anterior edge of the genital opening and is 1.83– 2.25mm in length, compared with M. norfolkensis (1.76–2.67mm). In the “ planiceps-beccarii-loriae complex” it is absent or less than 1.6mm in length.
M. eleryi is unique amongst Australian Mormopterus species by its possession of a glans penis with serrated lateral flanges ( Figure 5 View FIGURE 5 ).
M. eleryi differs from all other examined Australian Mormopterus species by its smaller skull size with GSL<14.41mm; CBL<13.64mm; MB<7.77mm; M3–M3<5.66m; C1–M3<5.23mm; SkHt<2.97mm; PZB<7.07mm; C1–C1<3.67mm while all other species exceed these values ( Table 3 View TABLE 3 ).
M. eleryi differs from all other Mormopterus species by its highly distinctive features of the upper molars (Figure 6). Within the “ planiceps-beccarii-loriae complex”, features of M1 and M2 are similar including a semi-isolated well-developed cone-shaped hypocone arising from the central body of the heel, and with the tip of the hypocone terminating close to the metaconule but separated via a short saddle. In M. eleryi , there is no typical hypocone, but rather a single crochet-hook shaped extension that arises from the disto-lingual edge of the heel. This structure does not appear to be homologous with the typical hypocone. The upper molars of M. eleryi also differ markedly from M. norfolkensis which itself has equally distinctive development of the heel region (described later).
M. eleryi differs from Malagasy M. jugularis Peters and Mascarene M. acetabulosus Hermann by the absence of a gular sac, the possession of four rather than six lower incisors, by the presence of genital projections, and by the shape of M1 and M2. M. eleryi differs from Sumatran M. doriae Andersen by the absence of a gular sac, the possession of four rather than six lower incisors. M. eleryi differs from Cuban M. minutus Miller by the absence of a gular sac, the presence of genital projections by the shape of M1 and M2. Differs from South American M. kalinowskii Thomas by the possession of P2, the shape of M1 and M2 but we have not examined other features. Differs from Peruvian M. phrudus Handley by the absence of a gular sac and smaller skull dimensions.
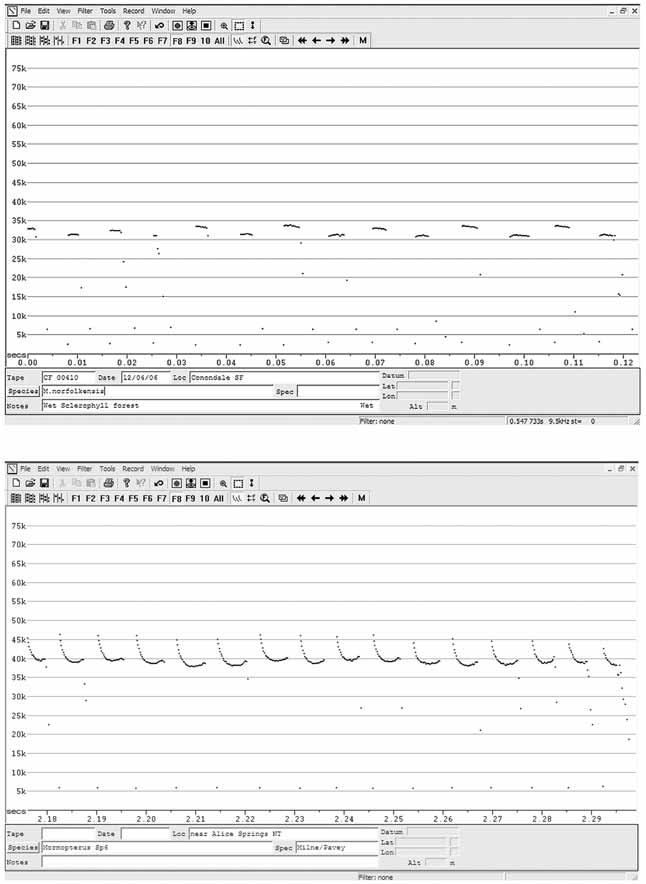
Echolocation call. The echolocation call differs from all Australian Mormopterus species calls by having strongly frequency modulated search phase pulses shaped like a reverse ‘J’ ( Figure 3 View FIGURE 3 ) compared with more flat shaped pulses of all other species. The call of M. eleryi has the highest average characteristic frequency (37kHz) compared with all other species of Australian Mormopterus (22–36kHz).
Sympatry. M. eleryi is sympatric with Species 1, 2, 3, 4 east, possibly 5 east, and occurs within 150km of M. norfolkensis .
Description. Externals. A small, gracile species, weighing between 5–8gms and forearm length 31.46– 35.92mm. Face with bristles as described above. Face lightly haired other than bristles and upper lips fringed. Vibrissae 3 to 4.4mm long, six each side of the muzzle. The muzzle much finer than other Australian Mormopterus species, face tapered to the large rhinarium, the margins of which are inflated to be proud of the muzzle. Nostril margins and median line of the septum are coarsely crenulated. The chin falls well short of the snout. There is no gular sac in either sex. Ears are triangular in shape and with the anterior edge straight. The ears are not joined across the forehead but separated by a gap 1.3–2.2mm. There is a small antitragus. The tragus is rounded.
The dorsal fur is bicoloured, hair shaft cream at the base for about 15% of the length and Vandyke Brown (121) to the tip. Hair shaft length is 4–5mm. Fur extends across the crown and tufts out onto the forehead through the margin of the ears. Ventral fur tricoloured, basal 10% cream, middle 70% Vandyke Brown (121) and tips cream. There appears to be some variation in pelage across geographic range. Northern specimens have slightly shorter and tighter groomed dorsal fur. In some southern specimens the dorsal fur appears a little less tightly groomed and tending to an overall appearance of more grey-brown than brown ( Figure 4 View FIGURE 4. M ).
Skin on ears, face, forearm, upper arm and legs is light coloured. The thumb pad is round. Toes each have three long hairs extending at least the length of the toes.
The glans penis is remarkable and completely unlike that of any other Australasian Mormopterus ( Figure 5 View FIGURE 5 ). The glans is not obviously differentiated from the penis shaft and in total is about 2.3mm in length. The glans is characterised by the presence of a serrated lateral flange that commences weakly from near the edge of the urethral opening and extends (becoming wider) to the shaft base. In profile the body of the penis is dorso-ventrally compressed and is acutely pointed at the tip.
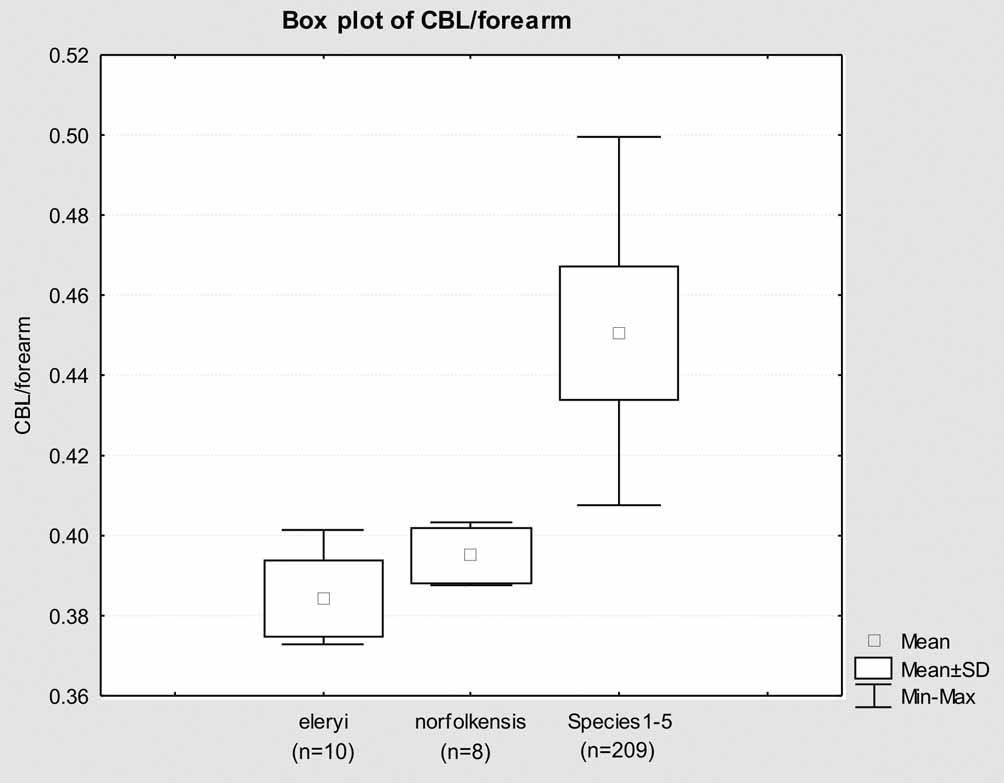
Skull and dentition. The skull ( Figure 7 View FIGURE 7 ) is small, both in absolute terms of most measures ( Table 3 View TABLE 3 ) and especially as a proportion of other body measures. For example, condylobasal length to forearm length averages 38.5% compared with 39.5% for norfolkensis and 45% for the " planiceps-beccarii-loriae complex" (Figure 8). The skull is more vaulted than in the " planiceps-beccarii-loriae complex".
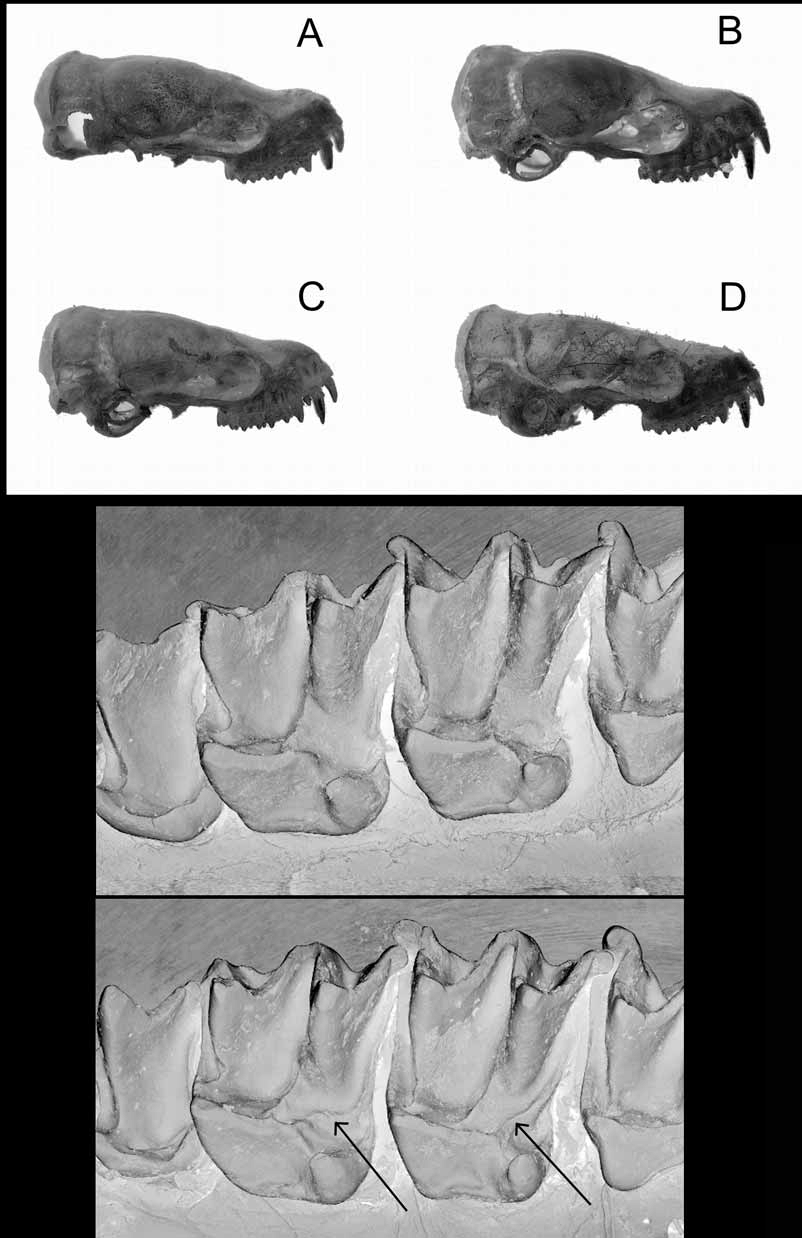
Our initial observation of the skulls of this species was based on the only four skulls that had been removed and these showed striking variation in the profile, with the three northeastern individuals having a distinct upwards inflection of the rostrum while the southwestern individual had no inflection. To determine whether this inflection was indicative of taxonomic distinction, age, location or gender, we x-rayed the eight available spirit specimens. Subsequently additional skulls were removed and genetic material from individuals with inflected and non-inflected skulls analysed. No allozyme differences were apparent between individuals of either skull type. Based on fourteen skull profiles (seven of each sex) we observed there was a range in the degree of inflection exhibited amongst individuals across the sampled distribution but the variation was not correlated to geographic location ( Figure 9 View FIGURE 9 ). There was no correlation between gender and inflection; for example the two Fletchervale individuals, a male and female, had the strongest inflection.
We had only eight skulls available to assess whether age was correlated with degree of inflection. We scored age according to the following three categories of tooth wear; none, intermediate and obviously well worn (canines and P4 generally showed little wear but wear in the upper molars was clearer and judged by the degree of wear of the post-protocrista) and compared these with two classes of magnitude of inflection; no or little inflection and marked inflection. Although these categories were somewhat subjective and sample size inadequate, there were some clear outcomes. For example, the two specimens from Fletchervale Station exhibited the strongest inflection; however, one was classed with no tooth wear while the other with wellworn teeth. Conversely, the specimens from Alcoota and Boomika represented the least inflection and had moderate and no tooth wear respectively. However, we regard this result with some caution because aging bats by tooth wear may be misleading. Our view is that the skull variation is not indicative of any taxonomic distinction.
The skull viewed dorsally shows the supraoccipital rounded, extending well back from the posterior margin of the lambdoidal crests. The development of the lambdoidal crests varies amongst specimens. In general they begin weakly at the base and either quickly diminish to no trace or continue as a small thickened ridge until they sweep markedly anteriorly where they meet the sagittal suture. There is either complete absence of a lambda or a small protuberance. A sagittal crest is very weak to absent. The interparietals and parietals are inflated, leaving a diamond shaped depression into the skull. There are distinct foramen (transverse sinus) in the suture between the parietal and interparietals. The mid-line of the skull is slightly concave from the lambda to well onto the rostrum. The postorbital and lachrymal processes are moderately developed, as is the supraorbital ridge. The posterior border of the narial emargination is round.
From a lateral view the skull is slightly sinuate, and as described above, variable in the degree of upward inflection of the rostrum near the posterior edge of the nasal emargination. Also seen from this view is the variable development of the lambdoidal crests and the lambda.
Dental formula: i1 /2 c 1/1 p2/ 2 m 3/3=30. The upper tooth row is not as crowded as in species of the “ planiceps-beccarii-loriae complex”. Although P2 is small, it sits in the line of the tooth row and its cingulum is the same height as, and touching, those of C1 and P4. As referred to previously, the striking feature of the dentition is the shape of M1 and M2. In addition to the crotchet hook shaped hypocone described earlier, M1 and M2 have a well-developed paraloph and metaloph ( Figure 9 View FIGURE 9 ). However the post protocrista in M1 and M2 appear to be quite variable amongst the specimens examined. For example, in the type specimen, there is a complete absence of this crest in both molars, while in the paratype SAM M15811, the crests are well developed ( Figure 9 View FIGURE 9 ). The lower dentition is shown in Figure 6.
Distribution, biology and conservation status. M. eleryi has now been recorded from 26 localities and exhibits an unusual distribution from central Australia (where the annual average rainfall is around 200mm) to eastern Australia including near Townsville (with 1100mm annual average rainfall) ( Figure 10 View FIGURE 10 ). Most records are from captures by Constantine traps or mist-nets set over pools or in the riparian zone of ephemeral creeks. Capture sites are usually associated with Eucalyptus camaldulensis and E. coolabah in the arid regions and with E. populnea , E. polyanthemos , E. melanophoia and Corymbia citriodora in woodlands and forest in the eastern part of the distribution. M. eleryi has been observed roosting in tree hollows ( Pennay 2006).
Little has been reported on the reproductive cycle. A female in late term pregnancy (single foetus) was captured in October in central Australia ( Churchill 1998). A maternity roost was noted in November 2005 in northern central New South Wales ( Pennay 2006). Churchill (1998) also notes that males had abdominal testes in January and October, but M. eleryi may have similar reproductive morphology as Species 4 in which the testes do not descend into scrotal sacs ( Krutzsch & Crichton 1987).
M. eleryi is not especially common in any region and generally accounts for less than 1% of individuals trapped or netted per night. M. eleryi (under the name Species 6) is listed as Endangered in New South Wales (Threatened Species Conservation Act 1995 No 101) and Vulnerable in South Australia (National Parks and Wildlife Act 1972 - 21.2.2008), but is not listed under any threatened category in the Northern Territory, Queensland or under federal legislation.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.