
Lophostoma occidentalis ( Davis & Carter, 1978 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.207288 |
|
DOI |
https://doi.org/10.5281/zenodo.5628094 |
|
persistent identifier |
https://treatment.plazi.org/id/03F8057A-FFF9-4F2D-90B5-FA38C1732704 |
|
treatment provided by |
Plazi |
|
scientific name |
Lophostoma occidentalis ( Davis & Carter, 1978 ) |
| status |
|
Lophostoma occidentalis ( Davis & Carter, 1978) View in CoL
Western Round-eared Bat
Figures 1 View FIGURE 1 and 5 View FIGURE 5
Tonatia silvicola occidentalis Davis & Carter, 1978: 6 View in CoL ; type locality “ 4 mi. W Suyo, 1000 ft., department of Piura, Perú.”
T [onatia]. silvicola occidentalis: Czaplewski, 1990: 235 View in CoL ; name combination.
Lophostoma aequatorialis Baker, Fonseca, Parish, Phillips, & Hoffmann, 2004 : 1; type locality “ Ecuador, Province of Esmeraldas, Estación Experimental La Chiquita, near San Lorenzo town (1°16’60”N, 78°49’60”W) (UTM zone 17: 748935 E 0 136902 N; 979 m).”
[ Lophostoma View in CoL ] occidentalis: Simmons, 2005: 406 View in CoL ; name combination.
L [ophostoma]. aequatorialis: Tirira, 2007: 276 ; name combination.
L [ophostoma]. s [ilvicolum]. occidentalis: Williams & Genoways, 2008: 270 View in CoL ; name combination.
Type material. The type material includes 18 specimens from three localities: ECUADOR: Loja, 15 mi N. of Catacocha, 2000 ft ( TCWC 11702 3); PERU: Piura, 4 mi. W. Suyo, 1000 ft ( TCWC 11703 Ƥ, 11704 Ƥ [holotype], 11705 Ƥ); PERU: Piura, Ayabaca, Paymas ( FMNH 81126 3); PERU: Morropon, Hacienda Bigote ( FMNH 81113 3, 81114–81119 Ƥ, 81120 3, 81121 Ƥ, 81122 3, 81123–81125 Ƥ). Davis & Carter (1978) assigned the locality of “ Peru: Piura: Salitral” to the 14 Peruvian specimens housed at the FMNH. After reviewing specimen labels, we affirm that these specimens came from the localities shown above.
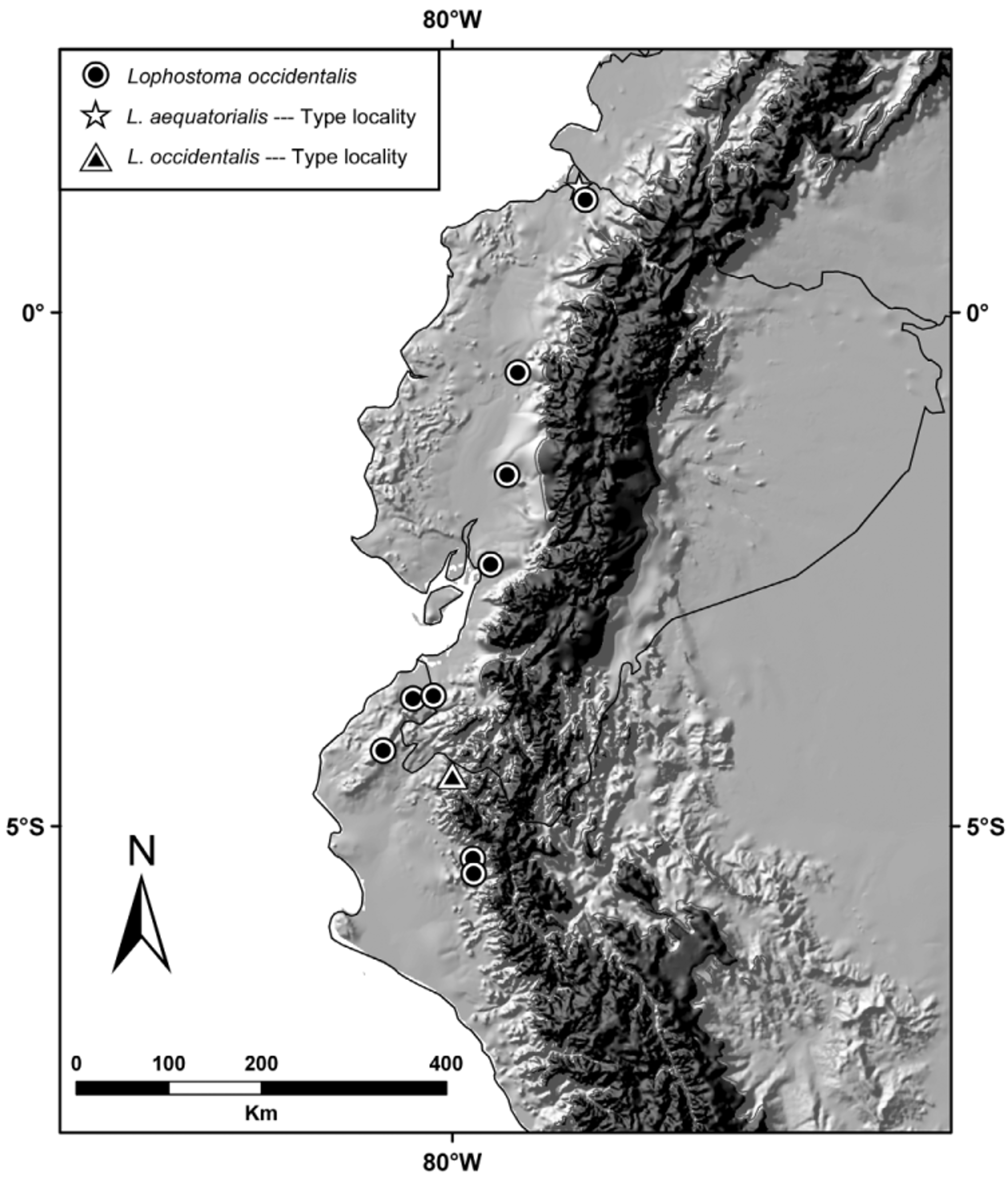
Distribution. Lophostoma occidentalis is known from the provinces of Esmeraldas, Los Ríos, and Pichincha in the western lowlands of Ecuador and from the departments of Tumbes and Piura in northeastern Peru ( Fig. 6 View FIGURE 6 ).
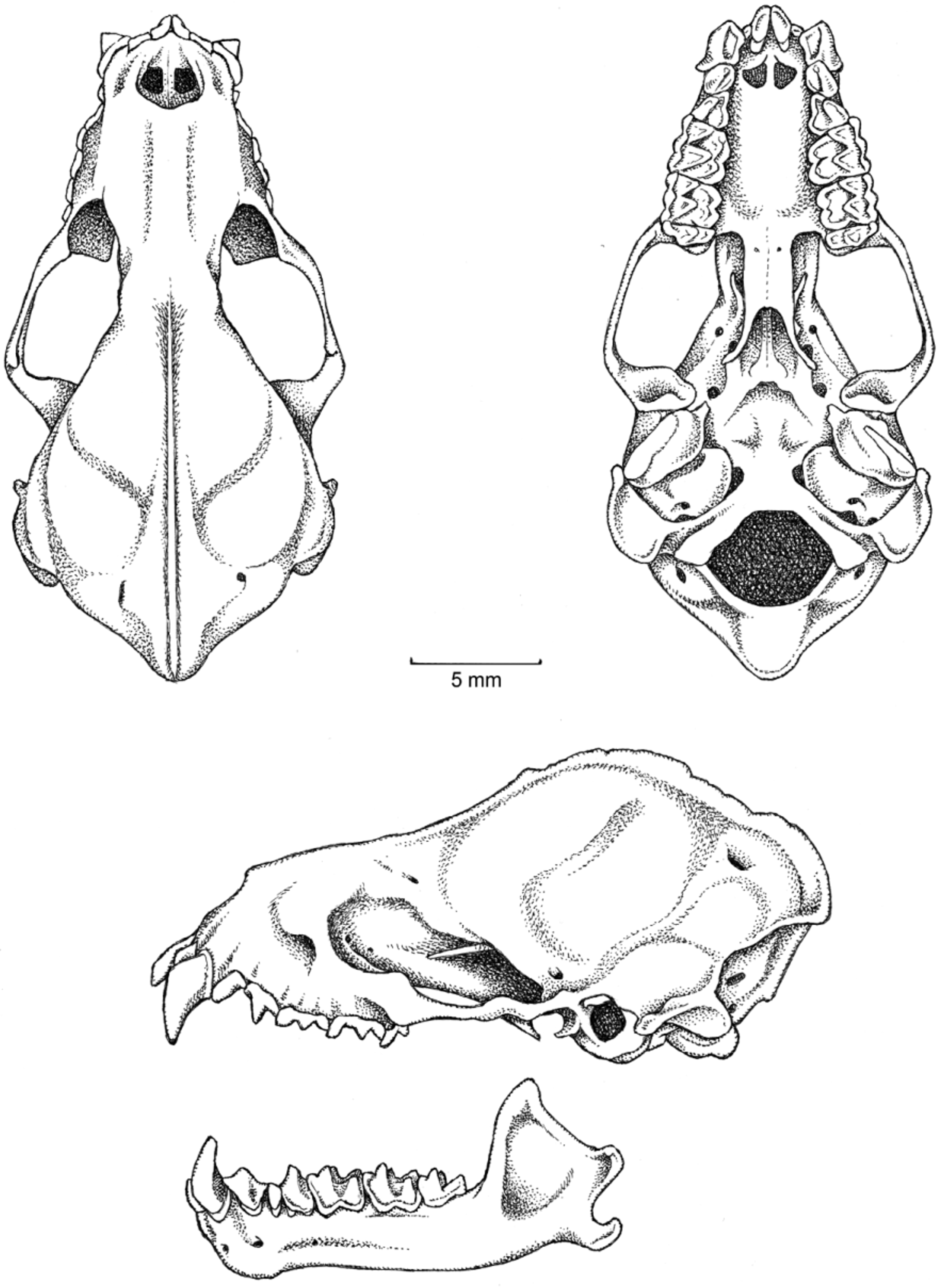
Emended diagnosis. Lophostoma occidentalis is a moderately large Lophostoma (FA 51.2–56.8, GLS 25.5– 28.7; CCL 21.5–23.7; Table 2), easily distinguished from L. brasiliense , L. carrikeri , and L. schulzi by its larger size and longer skull (Table 2), from L. schulzi and L. silvicolum by its paler throat, venter and auricular patches, and from L. yasuni by the restriction of white–pale ventral hairs to the throat and chest in L. occidentalis . L. occidentalis has white to pale post-auricular patches ( Fig. 1 View FIGURE 1 ) and the dorsal surface of the forearm is naked. Lateral projection of mastoid process is well developed ( Fig. 5 View FIGURE 5 ); basioccipital is wide; basisphenoid pits are shallow, weakly separated by a septum ( Fig. 5 View FIGURE 5 ); clinoid processes thin and curved laterally. Labial cingulum of the upper canines absent; P3 distal accesory cusp present; P4 distal accessory cusp present; P4 labial cingulum absent; M1 ectocingulum strongly developed; M1 mesostyle absent; M1 mesostylar crest present; M1 anterior ectoflexus deep and posterior ectoflexus shallow; M1 lingual cingulum present; M2 mesostyle absent; M2 mesostylar crest present; M2 lingual cingulum present.
Description and comparisons. Lophostoma occidentalis is not sympatric with any other species of Lophostoma . Lophostoma brasiliense is the smallest species of the genus and, with a forearm length less than 40 mm (Table 2), is unlikely to be confused with L. occidentalis , a moderately large Lophostoma (FA 51.2–56.8, GLS 25.5–28.7; CCL 21.5–23.7; Table 2). Linear measurements of L. occidentalis overlap with those of L. evotis , L. silvicolum , and L. yasuni ; whereas L. occidentalis is larger than L. brasiliense and L. schulzi (Table 2). Dorsal pelage in all Lophostoma species is dark brown and long, individual hairs tricolored with a short white base that makes up roughly 15% of the total length of each hair, a long dark brown (~80% of the total length of each hair) subterminal band, and a very short pale to whitish terminal band; gular fur is whitish in L. occidentalis , L. carrikeri , and L. yasuni (dark brown in L. brasiliense , L. evotis , L. schulzi , and L. silvicolum ); L. occidentalis and L. evotis present white to pale gray post-auricular patches (absent in L. brasiliense , L. carrikeri , L. schulzi , L. silvicolum , and L. yasuni ); post-auricular patches not connected by a thin line of pale hairs to the pale fur on the chest in L. occidentalis (connection present in L. evotis ); ventral fur white to beige across the chest but restricted laterally over the stomach and abdomen by the presence of pale brown fur on the side of the body in L. occidentalis , L. carrikeri , and L. yasuni (pale brown in L. brasiliense and L. schulzi ; pale to dark brown chest in L. evotis and L. silvicolum ); abdominal fur pale brown in L. occidentalis , L. brasiliense , and L. schulzi (white in L. carrikeri and L. yasuni ; dark brown in L. evotis and L. silvicolum ). Folds in the naked pinna are well marked; band of skin connects the internal base of the pinnae. Uropatagium sparsely haired on the proximal third and nearly naked posteriorly. The dorsal surface of the forearm is naked in L. occidentalis , L. schulzi , and L. silvicolum (the proximal third covered with sparse, short hair in L. brasiliense , L. carrikeri , L. evotis , and L. yasuni ); ventrally the proximal third of the forearm covered with long pale brown hair in L. occidentalis , L. brasiliense , L. carrikeri , and L. evotis (short pale brown hair in L. schulzi and L. silvicolum ). The dorsal surface of the forearm, digits, and legs, and sagittal midline of noseleaf lack wart-like granulations in L. occidentalis , L. brasiliense , L. carrikeri , L. evotis , and L. silvicolum (present in L. schulzi ). Metacarpal III is shorter than metacarpal V. Tibia is naked. Dorsal surfaces of the feet are naked; calcar longer than foot. L. occidentalis , L. brasiliense , and L. silvicolum have an elongated clitoris but the labia are not swollen; in L. carrikeri the clitoris is elongated and labia are swollen; whereas in L. schulzi the clitoris is remarkably elongated and resembles the penis of males. Two genal vibrissae are present in L. occidentalis ; along with approximately eight submental vibrissae on each side of chin and two interramal papillae each bearing several short vibrissae.
The skull of Lophostoma occidentalis has a slender rostrum with an accentuated postorbital constriction resembling that of L. brasiliense and L. carrikeri (rostrum is robust in L. evotis , L. schulzi , L. silvicolum , and L. yasuni ). Sagittal crest well developed in males (FMNH 81120) and moderately developed in females (FMNH 81119). Lateral projection of mastoid process is well developed in L. occidentalis , L. evotis , L. schulzi , L. silvicolum , and L. yasuni (absent or weakly developed in L. brasiliense ; moderately developed in L. carrikeri ). The basioccipital is wide; basisphenoid pits are shallow, weakly divided by a septum. Clinoid processes thin and curved laterally. The mandibular rami in L. occidentalis are variable; some individuals have curved rami (FMNH 81122), while others are straight (FMNH 81119, Fig. 5 View FIGURE 5 ).
Upper central incisors (I1) well developed and orthodont; outer upper incisors (I2) well developed. I2 displaced dorsally and excluded from occlusal toothrow by close apposition of the I1 and canines. Height of I1 greater than the height of I2 but less than twice the height of I 2 in L. occidentalis , L. schulzi , and L. silvicolum (height of I1 at least twice that of I 2 in L. brasiliense , L. carrikeri , and L. evotis ). Weakly developed indentation present on the lingual cingulum of the upper canine. Labial cingulum of the upper canines absent in L. occidentalis and L. brasiliense (weakly developed in L. carrikeri , L. evotis , and L. silvicolum ; strongly developed in L. schulzi ). P3 well developed; P3 postparacrista decreases sharply in height near paracone and does not extend to distal edge of tooth in L. occidentalis , L. silvicolum , and L. schulzi (postparacrista extends as a continuous crest to the distal aspect of the tooth in L. brasiliense , L. carrikeri , and L. evotis ); P3 distal accessory cusp present in L. occidentalis (absent in L. brasiliense , L. carrikeri , L. evotis , L. schulzi , and L. silvicolum ). P3 labial cingulum weakly developed in L. occidentalis , L. carrikeri , L. evotis , L. silvicolum , and L. schulzi (absent in L. brasiliense ). P4 postparacrista decreases sharply in height near paracone and does not extend to distal edge of tooth in L. occidentalis , L. silvicolum , and L. schulzi (P4 postparacrista extends as a continuous crest on the distal aspect of the tooth in L. brasiliense , L. carrikeri , and L. evotis ); P4 distal accessory cusp present in L. occidentalis (absent in L. brasiliense , L. carrikeri , L. evotis , L. schulzi , and L. silvicolum ). P4 length greater than width in occlusal view in L. occidentalis , L. brasiliense , L. carrikeri , L. evotis , and L. silvicolum (length and width subequal in L. schulzi ); P4 labial cingulum absent in L. occidentalis and L. brasiliense (weakly developed in L. carrikeri , L. evotis , L. schulzi , and L. silvicolum ). Metacone and paracone of M1 subequal in height; M1 ectocingulum strongly developed in L. occidentalis and L. silvicolum (weakly developed in L. brasiliense , L. carrikeri , and L. schulzi ; absent in L. evotis ); M1 postparacrista does not contact premetacrista, trigon open labially in L. occidentalis , L. evotis , and L. silvicolum (postparactista contacts premetacrista on labial aspect of M1, trigon closed labially in L. brasiliense , L. carrikeri , and L. schulzi ); M1 mesostyle absent in L. occidentalis (present in L. brasiliense , L. carrikeri , L. evotis , L. silvicolum , and L. schulzi ); M1 mesostylar crest present in L. occidentalis and L. silvicolum (absent in L. brasiliense , L. carrikeri , L. evotis , and L. schulzi ); M1 parastyle present in L. occidentalis , L. brasiliense , L. evotis , L. schulzi , and L. silvicolum (absent in L. carrikeri ); M1 anterior ectoflexus deep and posterior ectoflexus shallow in L. occidentalis and L. evotis (both deep in L. silvicolum ; both shallow in L. brasiliense , L. carrikeri , and L. schulzi ); M1 hypocone moderately to well developed in L. occidentalis , L. brasiliense , L. schulzi , and L. silvicolum (absent or poorly developed in L. carrikeri and L. evotis ); M1 lingual cingulum present in L. occidentalis (absent in L. brasiliense , L. carrikeri , L. evotis , L. schulzi , and L. silvicolum ). Metacone and paracone of M2 subequal in height; M2 ectocingulum strongly developed in L. occidentalis and L. silvicolum (weakly developed in L. brasiliense , L. carrikeri , and L. schulzi ; absent in L. evotis ); M2 postparacrista does not contact premetacrista, trigon open labially in L. occidentalis , L. evotis , and L. silvicolum (postparactista contacts premetacrista on labial aspect of M2, trigon closed labially in L. brasiliense , L. carrikeri , and L. schulzi ); M2 mesostyle absent in L. occidentalis (present in L. brasiliense , L. carrikeri , L. evotis , L. silvicolum , and L. schulzi ); M2 mesostylar crest present in L. occidentalis and L. silvicolum (absent in L. brasiliense , L. carrikeri , L. evotis , and L. schulzi ); M2 parastyle present in L. occidentalis , L. brasiliense , L. evotis , L. schulzi , and L. silvicolum (absent in L. carrikeri ); M2 hypocone moderately or well developed in L. occidentalis , L. brasiliense , L. evotis , L. schulzi , and L. silvicolum (absent in L. carrikeri ); M2 lingual cingulum present in L. occidentalis (absent in L. brasiliense , L. carrikeri , L. evotis , L. schulzi , and L. silvicolum ).
Crowns of the lower inner incisors are wider than long in L. occidentalis and L. silvicolum (subequal in L. brasiliense , L. carrikeri , and L. schulzi ; length greater than width in L. evotis ). Labial cingulum of the lower canines absent in L. occidentalis , L. brasiliense , and L. carrikeri (present in L. evotis , L. schulzi , and L. silvicolum ). p3 aligned in toothrow when seen in occlusal view in L. occidentalis , L. carrikeri , L. schulzi , and L. silvicolum (p3 labially displaced, not in line with other teeth in L. brasiliense and L. evotis ). Labial p4 cingulid straight in L. occidentalis , L. silvicolum , and L. schulzi (undulate in L. brasiliense , L. carrikeri , and L. evotis ).
Natural history. The northern populations of Lophostoma occidentalis inhabit evergreen lowland forest characterized by the presence of Brosimum utile , Castilla elastica (Moraceae) ; Wettinia quinaria , Phytelephas aequatorialis (Arecaceae) ; Guarea polymera (Meliaceae) ; Otoba gordoniiflora (Myristicaceae) ; Inga silanchensis (Fabaceace) ; Theobroma gileri (Malvaceae) ; and Xanthosoma daguense ( Araceae ) ( Gentry 1986, Céron et al. 1999). In contrast, the southern populations inhabit lowland deciduous forest characterized by the presence of Loxopterygium huasango (Anacardiaceae) ; Tabebuia chrysantha , Tecoma weberbaueriana (Bignoniaceae) ; Cochlospermum vitifolium (Bixaceae) ; Ceiba trichistandra , Eriotheca ruizii (Malvaceae) ; Cordia lutea , Cordia peruviana (Boraginaceae) ; Bursera graveolens (Burseraceae) ; Colicodendron scabridum (Capparaceae) ; Ipomoea carnea , Ipomoea philomega (Convolvulaceae) ; Muntingia calabura (Muntingiaceae) ; Acacia macracantha, Pithecellobium multiflorum, Prosopis pallida (Fabacea); Psittacanthus tumbecensis (Loranthaceae) ; and Ficus jacobii (Moraceae) ( Odar 2010, Linares-Palomino & Pennington 2007, Leal-Pinedo & Linares Palomino 2005, Linares- Palomino & Ponce-Alvarez 2005).
In Ecuador, Lophostoma occidentalis have been captured with Micronycteris megalotis , Micronycteris sp. A, Micronycteris sp. B, Mimon crenulatum , Phyllostomus elongatus , P. hastatus , Tonatia saurophila , Rhinophylla alethina , Carollia castanea , C. perspicillata , Artibeus aequatorialis , A. lituratus , Chiroderma trinitatum , Dermanura spp., Platyrrhinus dorsalis , P. matapalensis , Sturnira luisi , and Vampyressa thyone ( Baker et al. 2004) . In Peru, L. occidentalis have been captured with Pteronotus davyi , Micronycteris megalotis , Mimon crenulatum , Phyllostomus hastatus , P. discolor , Phylloderma stenops , Vampyrum spectrum , Desmodus rotundus , Diaemus youngi , Lonchophylla hesperia , Glossophaga soricina , Carollia brevicauda , C. perspicillata , Artibeus fraterculus , A. aequatorialis , A. lituratus , Chiroderma villosum , Platyrrhinus matapalensis , Sturnira luisi , Uroderma bilobatum , Eptesicus innoxius , Myotis nigricans , M. riparius , Rhogeessa velilla, Molossus molossus, Nyctinomops aurispinosus ( Pacheco et al. 2007, 2009).
A lactating female was captured in January ( Baker et al. 2004). Two bat flies Mastoptera minuta ( Costa Lima 1921) and Trichobius dybasi Wenzel 1966 have been found on L. occidentalis (J. Chavez, pers. comm.). Tricholeiperia peruensis Va rga s et al. 2009 was described as a new species of nematode from L. occidentalis captured at Cerros de Amotape National Park, Tumbes, Peru.
Karyology. The karyotype is 2 n = 34, FN = 62. The autosomes comprise a graded series of 15 pairs of metacentric and submetacentric chromosomes that range from large to small. No biarmed autosome consistently has an arm ratio less than one to two; however, in some spreads, pair 13 is subtelocentric in centromere placement. The smallest pair of autosomes is acrocentric. The X is a medium-sized submetacentric and the Y is a small acrocentric chromosome, approximately twice as large as the pair of acrocentric autosomes ( Baker et al. 2004).
Remarks. A subfossil left upper canine of Lophostoma occidentalis was reported from the Late Pleistocene (Lujanian) in the Talara tar seeps, Peru ( Czaplewski 1990). Myotis sp., Eptesicus cf. innoxius , and Eptesicus cf. fuscus were also recovered from the same deposits ( Cadenillas & Martinez 2005).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lophostoma occidentalis ( Davis & Carter, 1978 )
| Velazco, Paúl M. & Cadenillas, Richard 2011 |
Lophostoma
| Simmons 2005: 406 |
Tonatia silvicola occidentalis
| Davis 1978: 6 |