
Lichenopora hispida: Hincks, 1880: 473
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2002)270<0001:NABFTV>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03D1878C-1930-FFDD-FD52-C3E9FC14C5D3 |
|
treatment provided by |
Felipe |
|
scientific name |
Lichenopora hispida: Hincks, 1880: 473 |
| status |
|
Lichenopora hispida: Hincks, 1880: 473 View in CoL . Disporella hispida: Borg, 1944: 230 View in CoL . Hayward
and Ryland, 1985a: 128. Alvarez, 1992: 204;
1994c: 493.
Lichenopora mamillata Lagaaij, 1952: 181 .
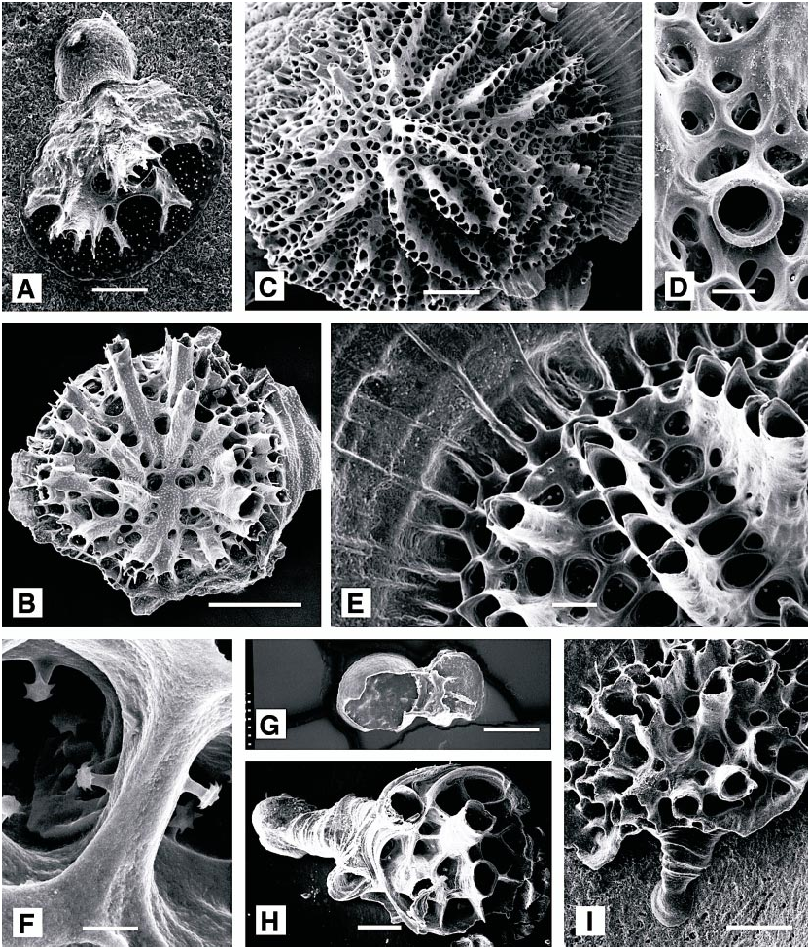
DESCRIPTION (AMNH 1043–1049; CMRR 2280–2282): Colonies yellow or white, multiserial, flat discoidal to low moundlike, commonly with slightly depressed center in infertile colonies, to at least 5.6 mm diameter; usually completely adnate with peripherally extended basal wall. Margin locally elevated where nearing contact with other encrusting organisms. Compound colonies reported elsewhere ( Hayward and Ryland, 1985a) not observed.
Autozooids budded only in continuous budding zone around colony perimeter. Ontogenetically older autozooids generally with isolated apertures aligned in single radiating rows, locally in short connate series. Younger zooids around perimeter of larger colonies generally in quincuncial pattern. Outer ends of ontogenetically oldest autozooids extend ed as peristomes, some with granules (more common on yellow colonies) or tiny spines (seen only on some white colonies) on upper surface. Small, blunt spines line the autozooidal chambers. Peristomes variable in length, generally short, diminishing in length toward perimeter, absent in youngest autozooids. Rim of peristomial apertures generally extended as paired spines on upper side, commonly with additional spines. Autozooidal apertures circular to oval, with long axis parallel with colony growth direction.
Autozooids separated by polygonal alveoli of equal to slightly smaller diameter. Ontogenetically older alveoli progressively filled centripetally, some closed entirely.
Brood chambers in central depressed area of colony, with domed roof of interior wall, extending as short lobes between proximal portions of inner portions of autozooidal rows. Ooeciostomes short, cylindrical, situated near perimeter of brood chambers; ooeciopores nearly circular, about 100 µm in diameter.
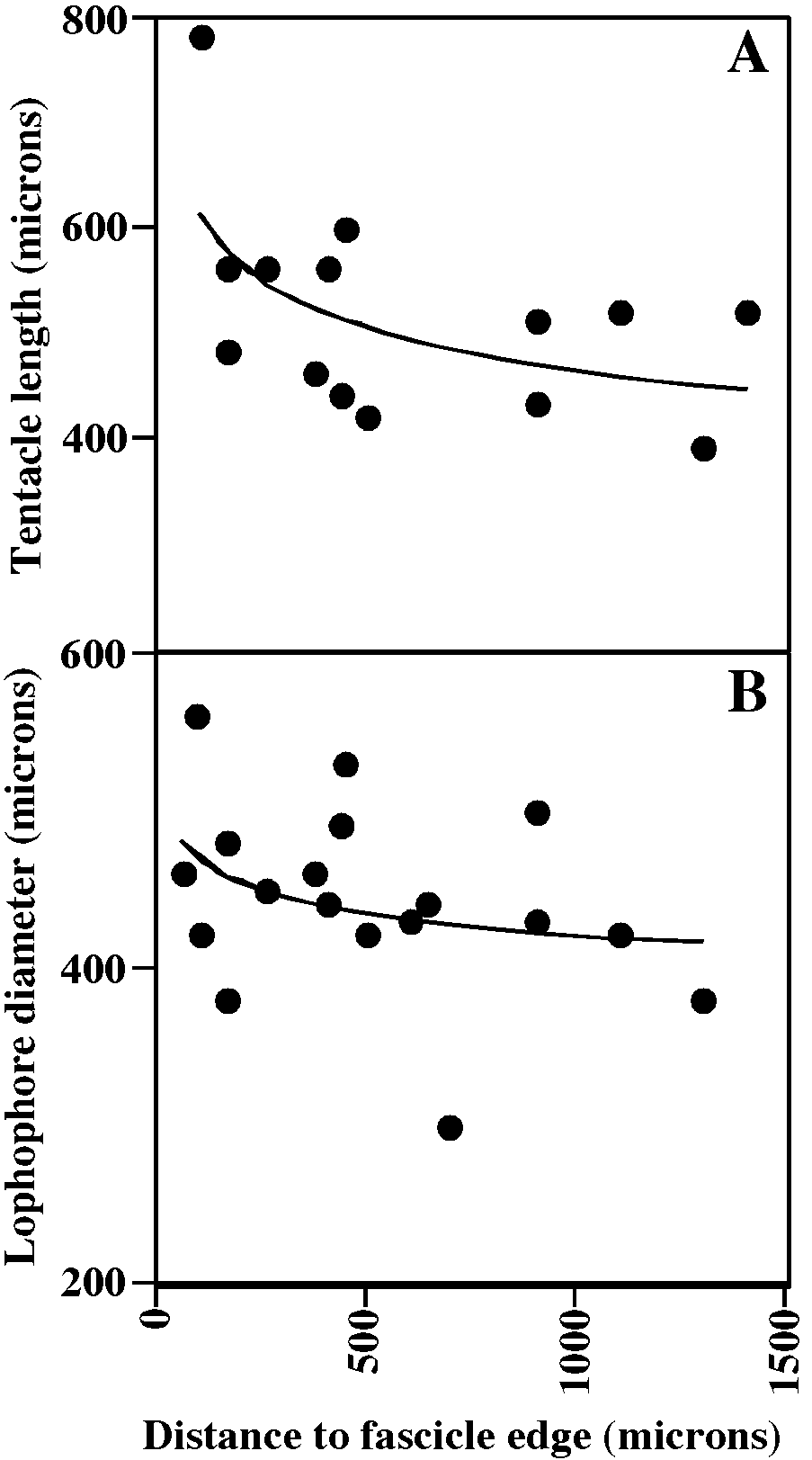
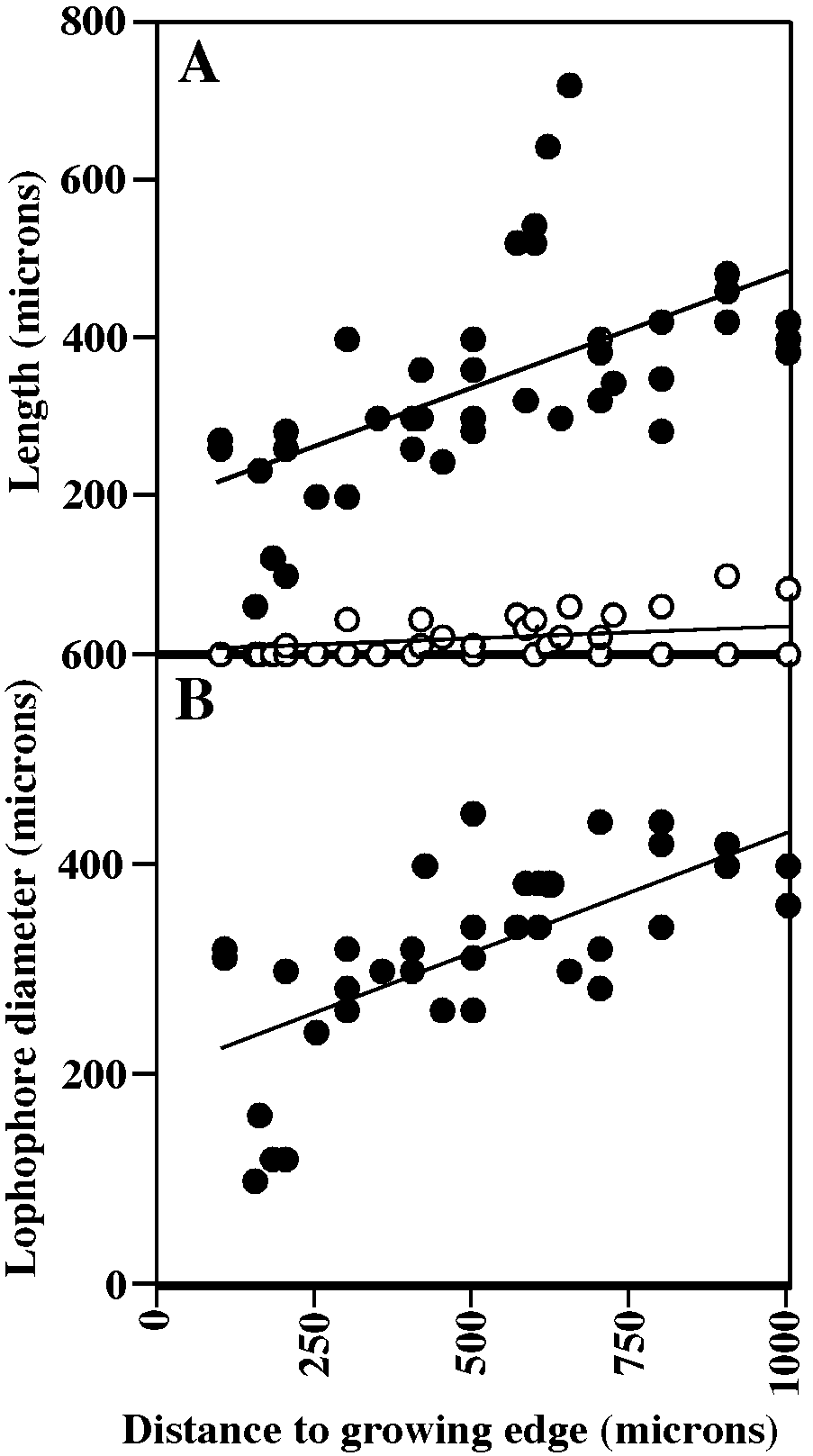
Tentacles clear, 9–10; lophophores obliquely truncate, broadly conical along colony margin to campylonemidan in ontogenetically older autozooids, increasing in size away from colony margin, youngest auto zooids equitentacled but progressively more inequitentacled with age; ontogenetically older autozooids commonly with lophophores elevated on introverts.
REMARKS: Several species names have been proposed for specimens of Disporella from the North Atlantic and Mediterranean, yet the differentiating characteristics of the proposed species are not clear. Alvarez (1992) attempted to sort out the species of Disporella from the region, naming four new species, and Gordon and Taylor (2001) have also addressed the taxonomic confusion in the genus. Although Alvarez (1992: 209) chose a neotype for D. hispida, Gordon and Taylor (2001) point out that there are two problems with the neotype: it is not topotypic, and its orifices are neither ‘‘expanded’’ nor are they hispid, which are attributes originally indicated for the species by Fleming (1828).
It is not clear whether one or two species are included here in ‘‘ Disporella hispida ’’. Four of the species described by Alvarez (1992) are hispid, i.e. have spines projecting from the skeletal aperture [ D. hispida (Fleming) ; D. robusta Alvarez, 1992 ; D. zurigneae Alvarez, 1992 ; D. alboranensis Alvarez, 1992 ]. Skeletally, the specimens from the Rovinj area fit only D. hispida among the six species of the genus that Alvarez (1992) described and illustrated (despite his choice of neotype, which was not illustrated). However, living colonies in our material fall into two discrete groups: yellow colonies with lophophores of 9 tentacles, and white colonies with lophophores of 10 tentacles. In ad dition, specimens from the Arctic that are identified as D. hispida by Shunatova and Ostrovsky (2001) have only 8 tentacles per lophophore, and none of the specimens have been seen with exposed introverts. More extensive documentation is needed in order to determine whether the various polypide morphologies of ‘‘ D. hispida ’’ are due to species differences or are ecophenotypes. However, both types of colonies are illustrated here. Fig. 58A–D View Fig portray yellow colonies; fig. 58E–J and 60A, B portray white colonies.
Brood chambers were found in some of the smallest colonies and were absent in the largest colonies, and where found, the colonies were fending off encroaching competitors for substratum space. This brood chamber occurrence pattern suggests that they are initiated when lethal environmental cues are encountered, previously suggested as common in encrusting cyclostomes ( McKinney and Taylor, 1997).
DISTRIBUTION: ‘‘ D. hispida ’’ occurs throughout the Mediterranean Sea, the Barents Sea, and on both shores of the northern Atlantic Ocean.
MEASUREMENTS (SKELETAL): ADMN 78 ± 10 µm, 60–100 (8, 100), ADMX 102 ± 12, 70–130 (7, 90), AS 211 ± 31, 140–300 (7, 90), AlvD 97 ± 18, 60–190 (7, 90), Gap 434 ± 62, 350–540 (1, 10). (POLYPIDE): IN 41 ± 60 µm, 0–220 (3, 49), LD 297 ± 56, 160– 440 (3, 46), MD 25 ± 5, 20–30 (2, 3), TLMn 292 ± 79, 120–420 (3, 31), TLMx 402 ± 132, 140–600 (3, 32).
GENUS PATINELLA GRAY, 1848 View in CoL
Patinella radiata ( Audouin, 1826) Figures 62C–I View Fig , 63 View Fig
Melobesia radiata Audouin, 1826: 235 .
Discoporella radiata: Busk, 1875: 32 . Waters, 1879: 276.
Lichenopora radiata: Hincks, 1880: 476 View in CoL . Borg, 1944: 222. Hayward and Ryland, 1985a: 124. Alvarez, 1990: 28, 1993: 262, 1994c: 500.
DESCRIPTION (AMNH 1048, 1050–1052; CMRR 2283): Colonies encrusting, multiserial, discoidal, small (up to 5.2 mm diameter in present material), with perimeter of calcified basal wall extending as a sheet beyond zooidal budding zone. Colonies basically white, some with purple granules below outer cuticle, giving light, speckled violet appearance to colonies viewed without magnification. Autozooids budded against basal wall in continuous zone around colony perimeter, rapidly diverging from basal wall at roughly 30° angle, commonly increased further away from basal wall. Zooids budded in linear series, generating singlezooid rows separated by alveoli, many of which also originate against the basal wall in the budding zone. Outer ends of zooids extended as peristomes, longer in ontogenetically older zooids, organized in connate fascicles. Small, blunt spines with stellate heads abundant in autozooidal chambers. In larger colonies, peristomes of younger zooids near colony perimeter possibly isolated rather than continuing the connate fascicles. Skeletal apertures of zooids slightly elongate parallel with growth direction and extended into acute projection on distal side.
Brood chambers located in central depression of colony that results from the radial and obliquely upward growth of autozooids. Brood chambers developing a convex roof covered by alveoli, with ooeciostome located centrally to peripherally within area defined by inner ends of autozooidal fascicles. Ooeciopores about 140 µm in diameter, circular to slightly oval; ooeciostomes short, and gently flared.
Protoecia exteriorwalled, low hemispherical,160–180 µm diameter, with low marginal rim. Ancestrula and initial asexually budded zooecia grow as an encrusting protoecial cone bounded by exterior wall, upper edge of which at some point bending back on itself, establishing radial growth of colony.
Tentacles clear, 8, shortest at colony margin (fig. 63A); lophophores campylonemidan, strongly obliquely truncate, grading from small and equitentacled along colony edge to largest and most inequitentacled adjacent to chimney at colony center (fig. 63B); lophophores closer to colony center support ed on short extroverts in some colonies (fig. 63A).
REMARKS: Living species previously included in Lichenopora Defrance (1823) apparently can no longer be considered to be part of this genus, which has as its type the Eocene species Lichenopora turbinata Defrance, 1823 (designated by d’Orbigny, 1853: 963). Gordon and Taylor (1997) revised the concept of the genus, restricting it to species which are either turbinate as is the type species, or have a short central pedunculate attachment with continued disklike expansion above the substratum. Both turbinate and pedunculate forms have large expanses of basal wall beyond the central point of attachment of the colony. The most recent known species of the Lichenopora , as redefined by Gordon and Taylor (1997), is Miocene. As suggested by Gordon and Taylor (1997), the generic name Patinella Gray, 1848 , with type species Madrepora verrucaria Linnaeus , is available for many living species formerly placed into Lichenopora . Patinell a and Disporella are here treated as distinct genera, based on Schäfer’s (1991) indication that the type species of the two genera ( Madrepora verrucaria and Disporella hispida Gray, 1848 ) differ in skeletal microstructure and in shape and perforation of the brood chambers.
DISTRIBUTION: The species occurs throughout the Mediterranean Sea, and ranges in the Atlantic Ocean from the Mediterranean northwards to southern Britain; reports from more distant regions are suspect ( Hayward and Ryland, 1985a).
MEASUREMENTS (SKELETAL): ADMN 83 ± 11 µm, 60–120 (5, 50), ADMX 110 ± 13, 80–140 (5, 50), Gap 215 ± 51, 100–340 (5, 46), GL 140–160 (2, 2), GW 120–160 (2, 2). (POLYPIDE): IH 15 ± 25 µm, 0–100 (3, 42), LD 324 ± 86, 100–450 (3, 41), MD 20 (1, 1), TLMn 193 ± 62, 60–320 (2, 26), TLMx 340 ± 133, 60–720 (3, 42).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lichenopora hispida: Hincks, 1880: 473
| HAYWARD, PETER J. & McKINNEY, FRANK K. 2002 |
Lichenopora mamillata
| Lagaaij, R. 1952: 181 |
Lichenopora hispida:
| Borg, F. 1944: 230 |
| Hincks, T. 1880: 473 |
Lichenopora radiata:
| Alvarez, J. A. 1994: 500 |
| Alvarez, J. A. 1993: 262 |
| Alvarez, J. A. 1990: 28 |
| Hayward, P. J. & J. S. Ryland 1985: 124 |
| Borg, F. 1944: 222 |
| Hincks, T. 1880: 476 |
Discoporella radiata:
| Waters, A. W. 1879: 276 |
| Busk, G. 1875: 32 |
Melobesia radiata
| Audouin, J. V. 1826: 235 |