
Arostrilepis beringiensis (Kontrimavichus & Smirnova, 1991) Gulyaev & Chechulin, 1997
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3608.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:32AAC94B-5793-4D51-8DCC-AF2D8AD5BCBD |
|
DOI |
https://doi.org/10.5281/zenodo.6147208 |
|
persistent identifier |
https://treatment.plazi.org/id/E07D87D9-FFF0-6519-68BA-F8DFBD2E8113 |
|
treatment provided by |
Plazi |
|
scientific name |
Arostrilepis beringiensis (Kontrimavichus & Smirnova, 1991) Gulyaev & Chechulin, 1997 |
| status |
|
Arostrilepis beringiensis (Kontrimavichus & Smirnova, 1991) Gulyaev & Chechulin, 1997
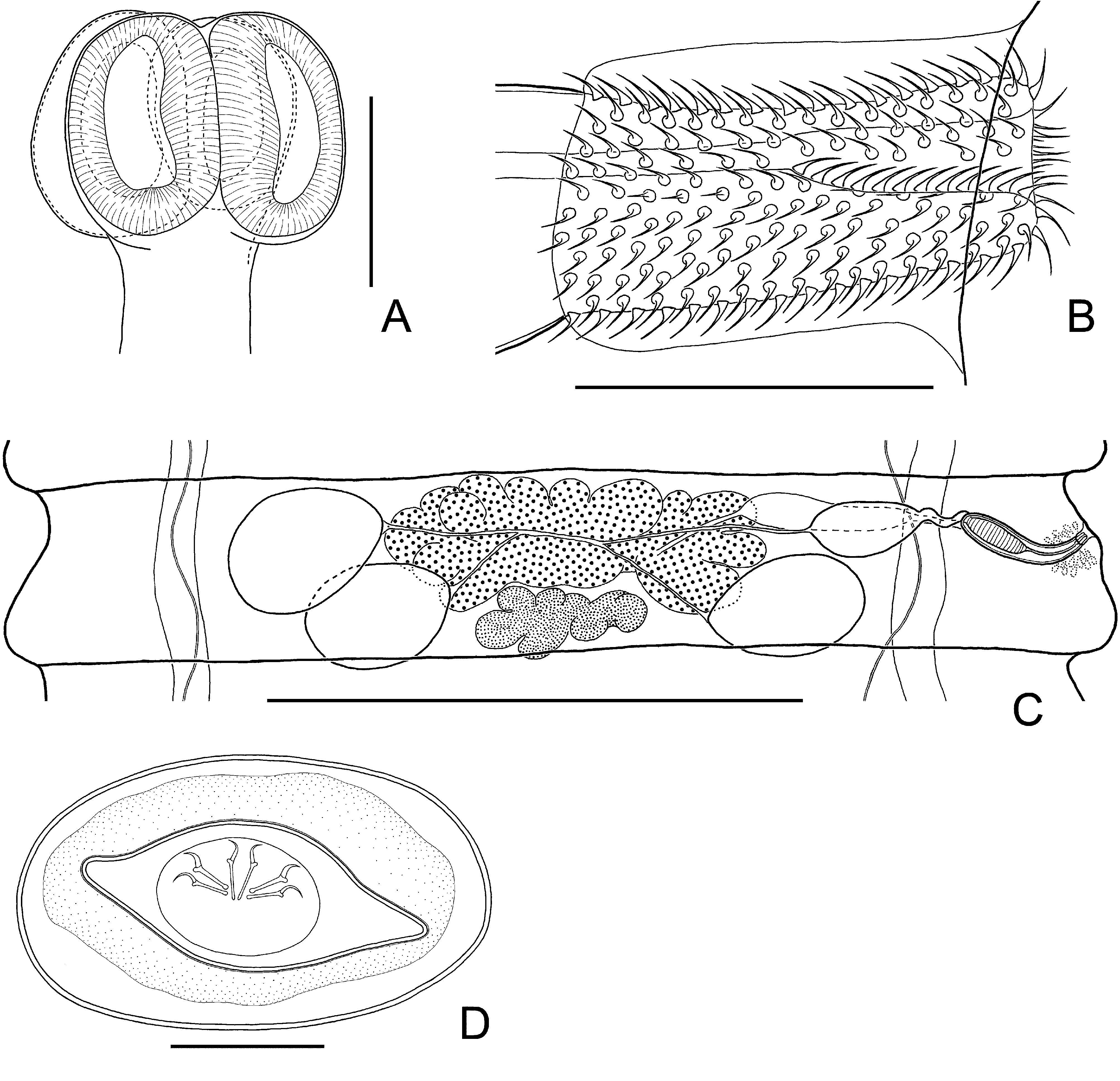
( Fig. 8 View FIGURE 8 )
Syn.: Hymenolepis beringiensis Kontrimavichus & Smirnova, 1991 .
Hosts in Nearctic: Lemmus trimucronatus (Richardson) and Synaptomys borealis (Richardson) .
Localities in Nearctic: Alaska- Barrow, Arctic Coastal plain; Brooks Range; eastern Beringia, Yukon Charley Rivers National Preserve.
Material examined: Vouchers include 4 specimens in Synaptomys borealis : MSB Para 1227, 1228, 1229 (field numbers AF 49480/ cyt- b sequence and 49480 A in Berlese’s; 49481/ cyt- b sequence, 2 specimens, multiple slides) by H. Henttonen et al., from the Yukon Charley Rivers National Preserve, Alaska across the Yukon River from Glen Creek Cabin (65o18’9”N, 142o02’14”W), 8 August 2001; MSB 1233, 1234 (AF 61601 B, 61601 C, 2 specimens) by H. Henttonen et al., from Gates of the Arctic National Preserve, Brooks Range, Alaska on unnamed lake off Koyukuk River (67o21’11”N, 150o51’00”W), 23 July 2002. Palearctic specimens: MSB 1235 (LEM 229a and 229b, 2 specimens) ex Lemmus sibiricus by V. Fedorov and G. Jarrell on the northwestern Taymyr Peninsula, Russia (ca., 75o34’N, 94o29’E), 24–26 June 1994; the specimen from this series (designated as LEM 117/ cyt- b sequence) from the northwest Taymyr was not available as a voucher. See Appendix 1 for listing of additional identified voucher specimens from the Nearctic and Palearctic.
Description: Based on 5 specimens. Fully developed strobila 145–196 mm long, with maximum width at pregravid or gravid proglottides, 1.5–1.8 mm. Scolex slightly compressed dorso-ventrally, 255–285 (270, n = 2) wide, clearly wider than neck. Suckers ovoid in surface view, 160–250 × 110–188 (198 × 149, n = 6), with thick walls, prominent ( Fig. 8 View FIGURE 8 A). Neck relatively long and narrow, 160–175 (170, n = 2) wide.
Dorsal osmoregulatory canals thin, 1–3 (2.1, n = 8) wide, situated predominantly in same sagittal plane as ventral canals. Ventral osmoregulatory canals 40–65 (54, n = 8) wide. Genital ducts may pass dorsally or between longitudinal osmoregulatory canals within same strobila; intersegmental variation without regularity. Development of proglottides gradual, protandrous.
Mature proglottides 160–200 × 825–1120 (174 × 980, n = 12), transversely elongate, trapeziform ( Fig. 8 View FIGURE 8 C). Testes usually 3 in number, almost of equal size, 140–195 × 100–140 (168 × 116, n = 15), round or oval, commonly situated in triangle with flat angle or rarely, testes form one row; poral testis separated from 2 antiporal testes by female gonads. Cirrus sac relatively short, 108–127 × 23–32 (117 × 27, n = 10), antiporal part of cirrus-sac commonly does not reach ventral longitudinal canal ( Fig. 8 View FIGURE 8 C). Genital atrium simple, cup-shaped, deep, opens laterally about middle of lateral proglottis margin, or slightly more anteriorly. Cirrus small, 36–47 × 9–12 (41 × 10, n = 10), cylindrical; armed along entire length with relatively small (up to 2.5–2.7 long) rosethorn-shaped spines ( Fig. 8 View FIGURE 8 B). Internal seminal vesicle, ovoid, 50–73 × 18–26 (63 × 21, n = 10), shorter than half of cirrus sac length ( Fig. 8 View FIGURE 8 C). External seminal vesicle 90–135 × 45–65 (111 × 52, n = 8), with size approximately equal to that of seminal receptacle.
Ovary 300–420 (365, n = 12) wide, median, fan-shaped, irregularly lobed, slightly overlapping testes ( Fig. 8 View FIGURE 8 C). Vitellarium 62–92× 152–225 (73 × 176, n = 10), postovarian, median, scarcely lobed. Vagina tubular, clearly distinct from seminal receptacle; ventral to cirrus sac. Seminal receptacle relatively small, 87–115 × 30–40 (101 × 36, n = 5).
Gravid proglottides 320–430 × 1300–1800 (366 × 1547, n = 8). Fully developed uterus labyrinthine, occupying entire median field and extending bilaterally beyond longitudinal osmoregulatory canals. Eggs 33–37 × 56–62, elliptical, with thin outer coat; oncosphere 12–15 × 18–21 ( Fig. 8 View FIGURE 8 D). Embryophore fusiform, 14–19 × 40–46, with straight polar processes. Embryonic hooks small, 7–8 long.
Remarks: Results of the present morphological analysis of specimens of A. beringiensis do not indicate a high degree of differentiation between cestodes from the Nearctic and Palearctic currently isolated across the Bering Strait (Makarikov & Kontrimavichus 2011). We observed that in specimens of A. beringiensis from the Nearctic the dimensions of the eggs, embryophores and oncospheres are larger relative to those from the Palearctic. We assume that the differences in egg measurements of the specimens are associated with the methods of specimen preparation rather than with geographical divergence of two populations. Eggs of A. beringiensis from the Palearctic were mostly compressed because these specimens, stained in Ehrlich’s haematoxylin, had been mounted permanently in Canada balsam. In contrast, gravid proglottides of specimens from the Nearctic were mounted in Berlese’s medium and as a consequence the eggs were not influenced by compression. Otherwise specimens we examined representing populations of A. beringiensis in bog lemmings and Nearctic brown lemmings from North America and Siberian brown lemmings from Eurasia do not appear to be differentiated morphologically relative to host species or specific geographic localities (Makarikov & Kontrimavichus 2011).
These are the first confirmed records of A. beringiensis in eastern Beringia and the Nearctic. Additionally, cestodes reported as H. horrida in L. trimucronatus (Nearctic brown lemming) from Okpilak River, Alaska (70o08’N, 143o38’W) by Haukisalmi and Henttonen (2001) (USNPC 89245) and those from Barrow, Alaska by R.L. Rausch (RLR 29831) are referable to this species. At Siberian localities including Lopatka Peninsula, Taymyr Peninsula, New Siberian Islands, and Wrangel Island, we confirm the presence of A. beringiensis , and redetermine records and specimens (USNPC 89243 and 89244) attributed to H. horrida by Haukisalmi and Henttonen (2001) at these sites. In Eurasia most records refer to cestodes in L. sibiricus , although other species including L. amurensis Vinogradov from eastern Siberia between the Lena and Kolyma Rivers and on the New Siberian Archipelago and L. portenkoi Tchernyavsky on Wrangel Island may be represented as hosts of this species (see Musser & Carleton 2005).
Arostrilepis beringiensis appears to be geographically widespread in lemmings (both Synaptomys and Lemmus ) at high latitudes in North America and in lemmings of Eurasia, although an association with species of Dicrostonyx Gloger requires confirmation (E.P. Hoberg, K.E. Galbreath, unpublished data; Haukisalmi & Henttonen 2001; Makarikov & Kontrimavichus 2011). Specimens attributed to Hymenolepis horrida have been reported as common parasites in Nearctic collared lemmings ( Dicrostonyx groenlandicus (Traill)) from near Barrow, Alaska (Kuns & Rausch 1950; Schiller 1952) and are also known in the Palearctic collared lemming (D.
torquatus Pallas) from northern Eurasia (e.g. Ryzhikov et al. 1978); in either case, these specimens were not available for study. Interestingly, Dicrostonyx (tribe Dicrostonychini ) is not considered phylogenetically close to the true lemmings ( Lemmini ) (reviewed in Musser & Carleton 2005), and these arvicolines may be derived from the earliest radiation of the subfamily linking Eurasia and North America and extending to 5–6 MYA (e.g. Conroy & Cook 1999). Such suggests that there is no particular basis to predict that the host distribution for A. beringiensis may extend to species of Dicrostonyx , unless it is present due to host colonization.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.