
Karnimata fejfari, Kimura & Flynn & Jacobs, 2017
|
publication ID |
https://doi.org/ 10.2478/if-2017-0011 |
|
persistent identifier |
https://treatment.plazi.org/id/8827BB0C-FF81-E205-E1F9-FC2AE3EBFB31 |
|
treatment provided by |
Diego |
|
scientific name |
Karnimata fejfari |
| status |
sp. nov. |
Karnimata fejfari sp. nov.
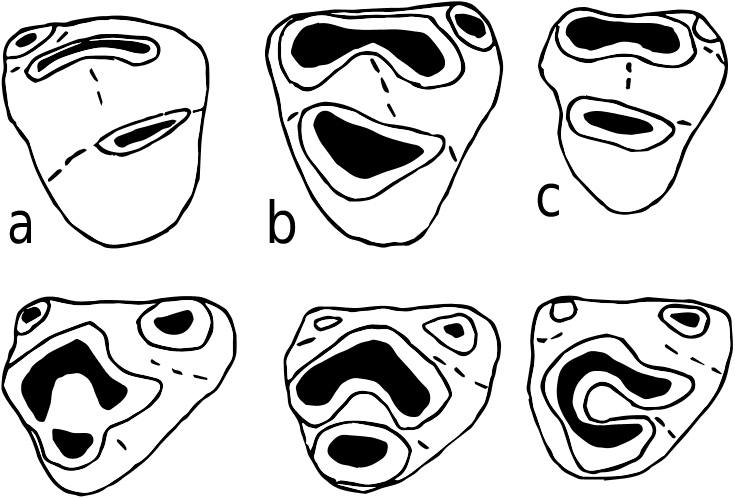
H o l o t y p e. YGSP 34546, left M1, Text-fig. 4c. View Text-fig
E t y m o l o g y. For Oldřich Fejfar, vertebrate paleontologist well known for his seminal works on Neogene mammal research, especially small mammals.
Ty p e l o c a l i t y. Y 311, upper part of the Nagri Formation, Siwalik Group, Chinji and Sethi-Nagri area of the Potwar Plateau, northern Pakistan, age 10.1 Ma.
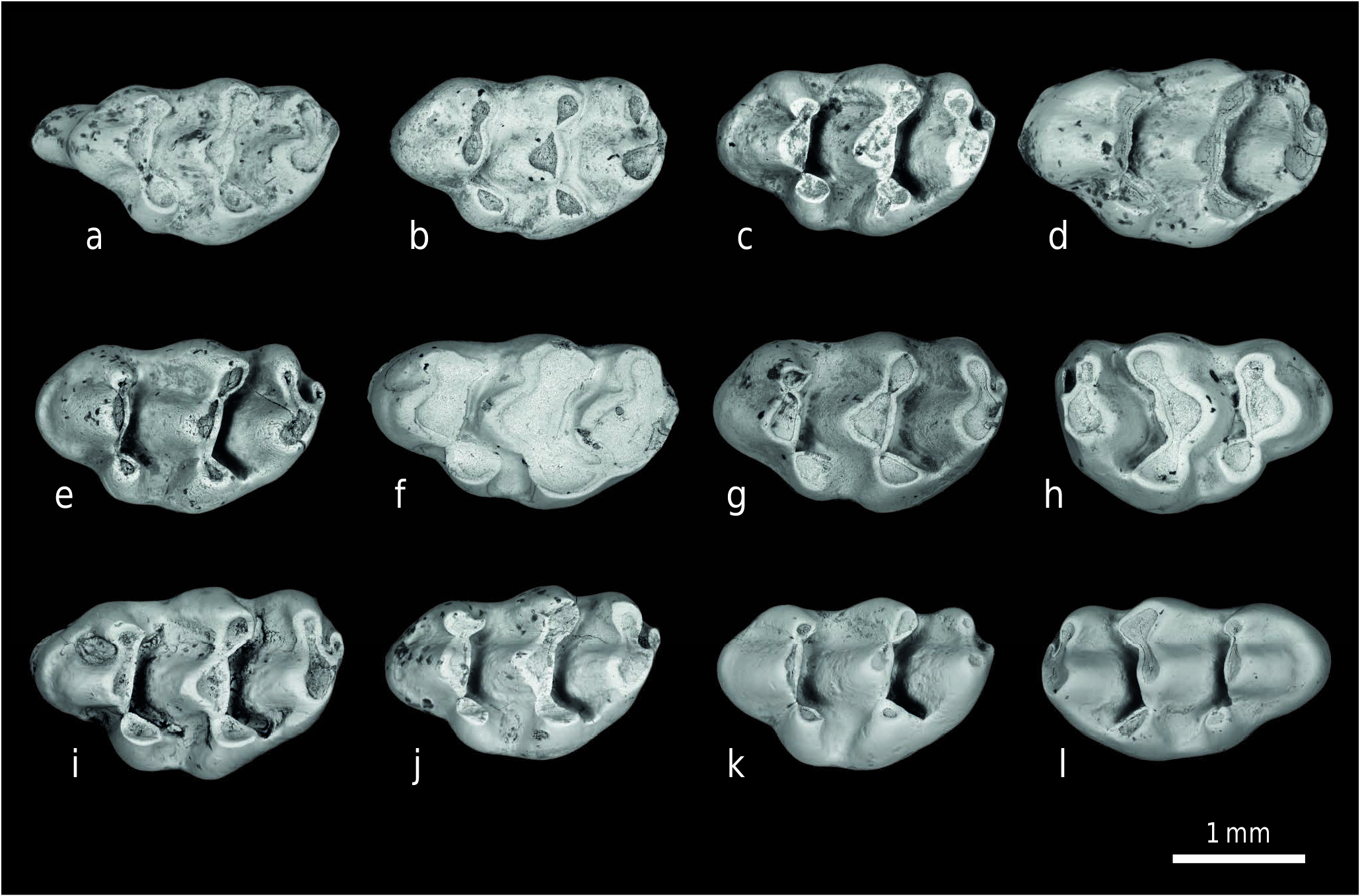
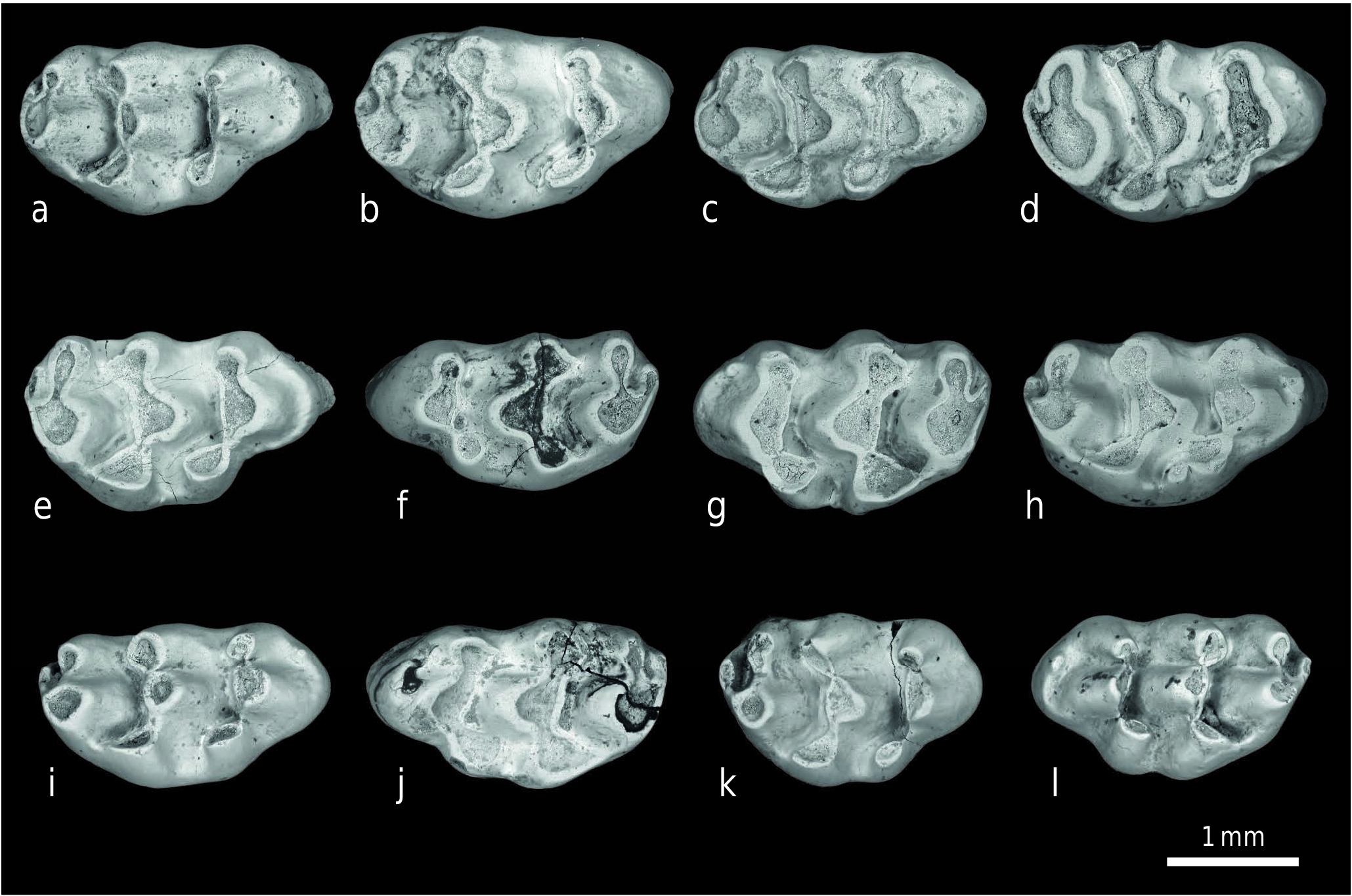
M a t e r i a l. 98 isolated molars from locality Y 311 (enumerated in Appendix). Additional material from other localities: 23 molars from Y 450, 161 molars from Y 259 ( Text-figs 4 View Text-fig , 5 View Text-fig ).
M e a s u r e m e n t s. See Appendix.
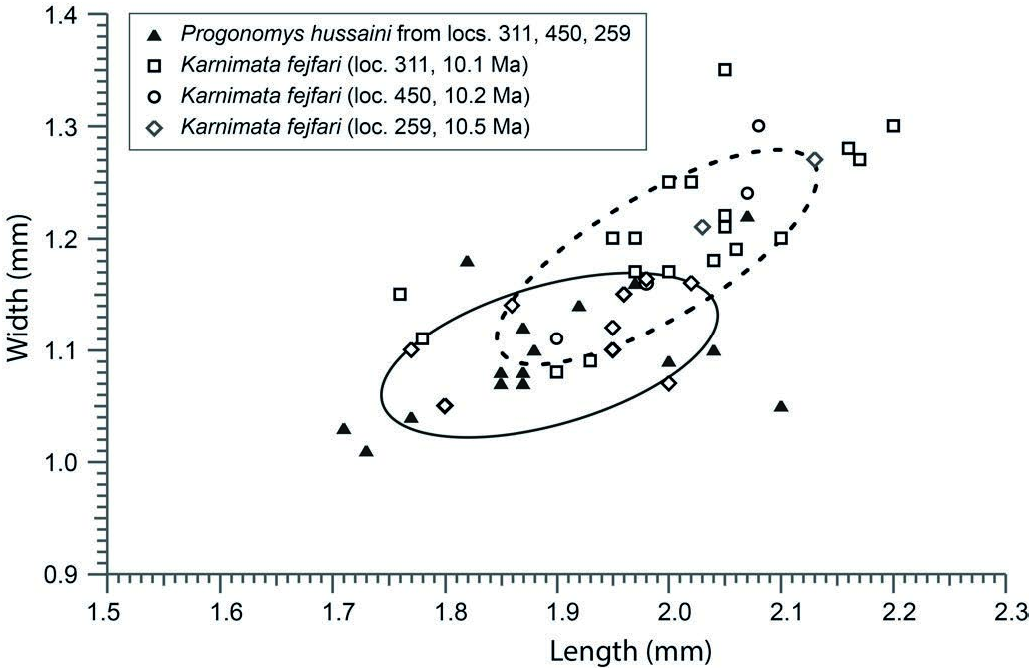
D i a g n o s i s. Karnimata slightly smaller than K. darwini , with anterostyle somewhat less anterior in position. M1 (L/W ratio = 1.65) more slender than K. darwini (L/W ratio = 1.56).
D e s c r i p t i o n. Molar cusp inclination is less than that of Progonomys hussaini , so that upper molar chevrons appear to lie relatively forward, even in early wear. M1 has eight major cusps, three in each of the first two chevrons. Its length/width ratio is 1.65. In the first chevron, the lingual anterocone is largest, labial anterocone smaller but prominent. The anterostyle (t1), generally rounded to somewhat laterally compressed, is linked to the posterolingual corner of the lingual anterocone, and is anterolingual to the protocone. The labial anterocone is more anterior in position than t1, and is rounded or occasionally shows a short posterior spur. In the second chevron, the protocone is the largest cusp, followed closely by the enterostyle (t4) and paracone. The enterostyle has a prominent posterior spur joining the hypocone low on its base. The paracone is completely isolated from the metacone (no hint of stephanodonty), although late in wear a short posterior spur may become evident. A short posterior cingulum is well developed and the precingulum, although variable, is normally present. M1 has an anteroposteriorly expanded lingual root, a large anterior root, and a posterior root. While roots are broken on many specimens, most but not all of the well-preserved teeth show an additional central rootlet.
M2 resembles M 1 in that it is robust with round cusps, not greatly inclined. The major chevron (paracone-protoconeenterostyle) dominates the tooth, but the anterostyle is a prominent cusp located anterolingually. The small labial anterocone is always present and usually shows wear. The enterostyle has a posterior spur in contact with the base of the hypocone. The metacone is small and separated from the paracone until late wear. The short posterior cingulum is usually clear. M2 has three roots.
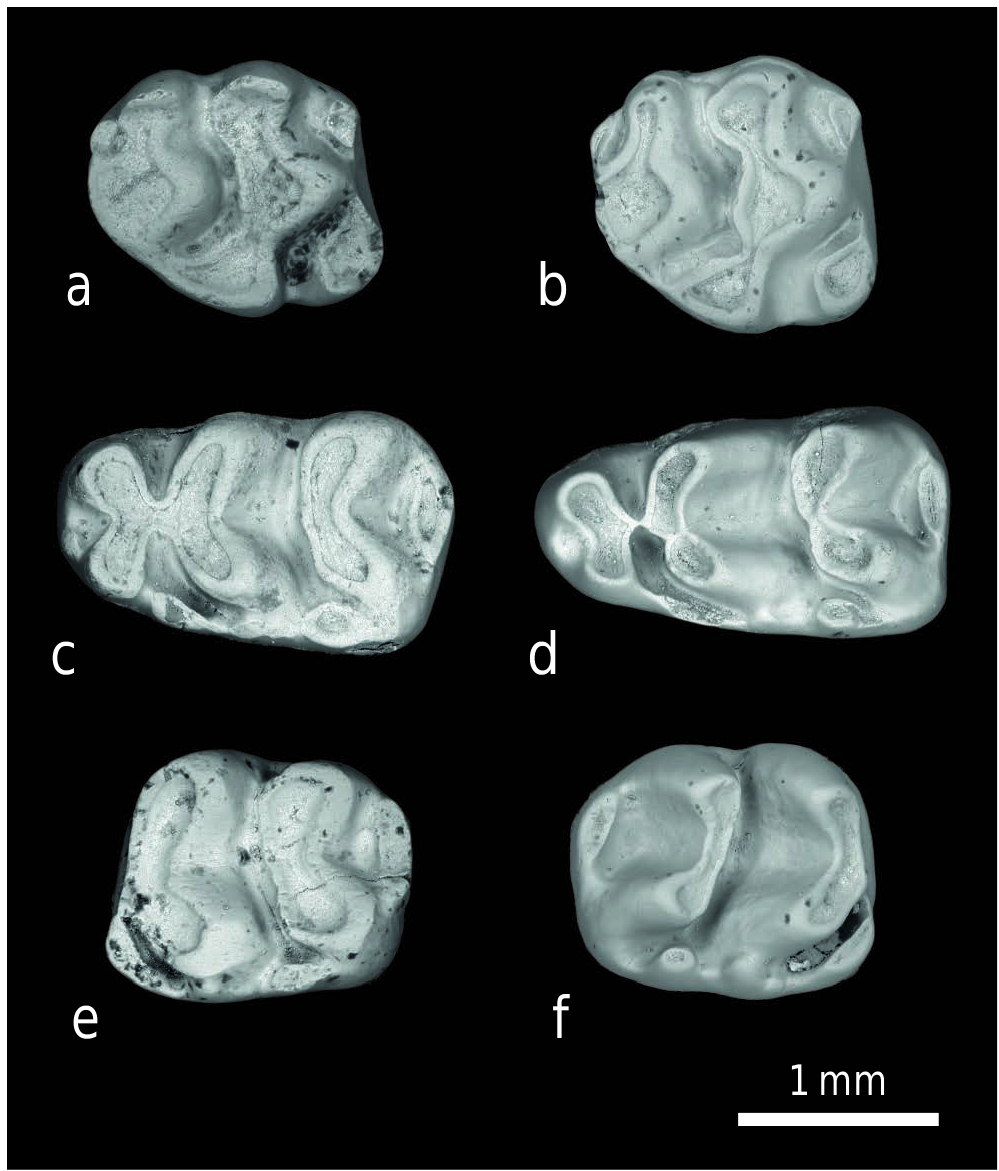
We found no convincing bimodal distinction among the M3 from the type locality Y 311 to assign them with confidence to K. fejfari versus Progonomys . The available specimens show some variation in size and possibly the larger ones represent mostly K. fejfari ; the smaller ones Progonomys hussaini . Also, the larger M3 generally have widths greater than lengths. All M3 have large anterostyles, but the labial anterocone differs in prominence among specimens; in some it is quite small. We propose without conviction that the larger specimens with strong labial anterocones represent K. fejfari . Three are listed in the appendix. One slightly smaller M3 from Y 450 (Appendix) has a strong labial anterocone and may also represent K. fejfari .
Lower molars are robust. The protoconid-metaconid and hypoconid-entoconid pairs of cusps are angled somewhat and span m1 broadly. The twinned anteroconid cusps are smaller, and there is generally a narrow shelf anterior to them, on which a minute medial anteroconid occurs in a few specimens. Wear on the posterior cingulum produces an oval loop of enamel. The labial cingulum is generally well developed, and usually bears accessory cusp c1, variably c3. Two roots anchor the tooth, and some specimens show a central rootlet. Posterior lower molars carry on the theme of m1 robustness. Their opposite cusp pairs are broad. The outline of m2 departs from rectangularity because its extended labial anteroconid supplies greater length than on the lingual side of the tooth. The hypoconid-entoconid cusp pair is nearly as broad as the protoconid-metaconid pair. The posterior cingulum is distinct. The labial cingulum is welldeveloped, bears c1, and usually has a cusp in position of c3. There are two roots. The m3 (see Cheema et al. 2000: fig. 6c) is distinguished by breadth of its anterior cusp pair to match the adjacent m2. The m3 is a small tooth with single, teardrop posterior cusp located somewhat lingual to the tooth midline. There is a small labial anteroconid, high enough to develop a wear facet. The labial cingulum is absent, except for an infrequent swelling of enamel representing c1. There is no posterior cingulum and two roots are present.
D i s c u s s i o n. In Siwalik murid assemblages of around 9 Ma (Dhok Pathan Formation, which overlies the Nagri Formation), two common species are clearly evident. Karnimata darwini , somewhat larger than K. fejfari , is the most abundant, followed by Progonomys debruijni , which is considerably smaller than P. hussaini . Nagri localities Y 311, Y 450, and Y 259 have the same two lineages, but the species are less derived, and they are close in size at those sites ( Text-fig. 6 View Text-fig ). Although size range overlaps with P. hussaini , K. fejfari is moderately larger with relatively broader, more heavily built molars. The morphological and size distinctions between the two lineages, so clear at 9 Ma, are subtle at 10 Ma.
Samples of isolated teeth from later Nagri localities currently available to us allow us to distinguish Karnimata from Progonomys for most teeth at most loci. There is some overlap in morphology such that some teeth may be misidentified. In fact, several specimens were assigned incorrectly to taxa in Kimura et al. (2013a, b, 2016). Following are the traits that we used to distinguish the species.
For M1, we utilized the defining characteristic of anterostyle position. This is not always absolutely distinctive for populations of 10.5 to 10 Ma. By 9.2 Ma, anterostyle position is a fixed diagnostic (= qualitative) character delimiting the separate species in their diverging lineages, but not so at 10 Ma ( Kimura et al. 2016). Progonomys has a more posterior anterostyle which tends to be laterally compressed, unlike the anterior, conical anterostyle of Karnimata . Progonomys hussaini has slightly more inclined cusps than Karnimata fejfari . This is not always clearly the case, especially in worn teeth, but is an additional aid in identification. As noted above, the double anterocone of Progonomys is more asymmetrically placed than is the M1 of Karnimata , with larger lingual anterocone and more asymmetrical anterior margin.
Most M2 of Karnimata fejfari are recognizable by moderate cusp inclination less than that of Progonomys , which is most evident for the protocone, the major anterocentral cusp. Its apex is not strongly inclined, such that the anterior surface of the cusp is steep ( Text-fig. 7 View Text-fig ). M2 is also relatively broader than in Progonomys , with a heavier lingual anterocone (t3).
Lower molars of Karnimata fejfari are more heavily built than those of Progonomys hussaini . The first molar has a strong labial cingulum, usually with prominent accessory cusps, and the double anteroconids are more divergent and independent. Lower m2 resembles m 1 in its strong labial cingulum and tooth breadth. The breadth comparable to that of m1 is apparent posteriorly as well as anteriorly with the second loph equal almost to the first loph. The broad posterior end of m2 is reflected in m3, whose anterior loph is broader than that of P. hussaini ( Text-fig. 8c View Text-fig ). The m3 of P. hussaini is smaller overall, and with a lower labial anteroconid. The contrast in m3 is seen in Cheema et al. (2000), in which their figures 6b and 6c represent P. hussaini and K. fejfari , respectively.
Among the third molars of Text-fig. 8 View Text-fig we illustrate several M3, which generally present difficulties for identification. Whereas the m3 locus shows two morphological states and sizes that are useful for species assignment, the morphological variation in M3 is more graded. Text-fig. 8e and 8f View Text-fig shows larger M 3 specimens with stronger anterostyle and labial anterocone, which we propose may represent K. fejfari . These teeth, and the somewhat smaller referred specimen from Y 450 ( Text-fig. 8g View Text-fig ) are wider than long. All three contrast with YGSP 34574 from Y 311 ( Text-fig. 8h View Text-fig ), which we attribute to P. hussaini . The much smaller M3 of Progonomys morganae ( Text-fig. 8d View Text-fig ) has low anterostyle and labial anterocone.
Our current interpretation sees Karnimata as a longlived lineage with roots in a diversification of Progonomys - like murids, perhaps beginning around 11 Ma. Karnimata represents the earliest clear indication of a major split in murid evolution, lying at the base of the Mus / Arvicanthis split ( Kimura et al. 2015). Early Karnimata as old as ~11 Ma is not easily distinguished from early Progonomys . Closer to 10 Ma, Karnimata fejfari encompasses a recognizable, early step in arvicanthine evolution. Karnimata survived to the end of the Miocene in the Indian subcontinent, and to the early Pliocene of Afghanistan ( Brandy 1979). Karnimata has been thought to occur in the late Miocene of Shanxi and Inner Mongolia, China, but Qiu and Li (2016) distinguish this material as new genus Karnimatoides.
Mein et al. (1993) considered the possibility that the holotype of Siwalik Karnimata darwini might represent the European species, Progonomys woelferi BACHMEYER et 1 mm
e f g h
WILSON, 1979, possibly rendering the genus name a junior synonym. However, Storch and Ni (2002) ruled this out, noting the anterior position of the anterostyle, which unlike Progonomys is characteristic of Karnimata . Kimura et al. (2013b, 2015) further showed that K. darwini is encased in a lineage separate from Progonomys , and that Karnimata is the most basal genus known of the tribes Arvicanthini - Otomyini-Millardini clade.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.