
Jassastaudei Conlan, 1990
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4939.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:F33F42D0-A139-4CE3-97D7-1314C12CF86B |
|
DOI |
https://doi.org/10.5281/zenodo.4580585 |
|
persistent identifier |
https://treatment.plazi.org/id/03B487DA-FFC4-D964-C9C8-18B5FD38F814 |
|
treatment provided by |
Plazi |
|
scientific name |
Jassastaudei Conlan, 1990 |
| status |
|
Jassastaudei Conlan, 1990 View in CoL
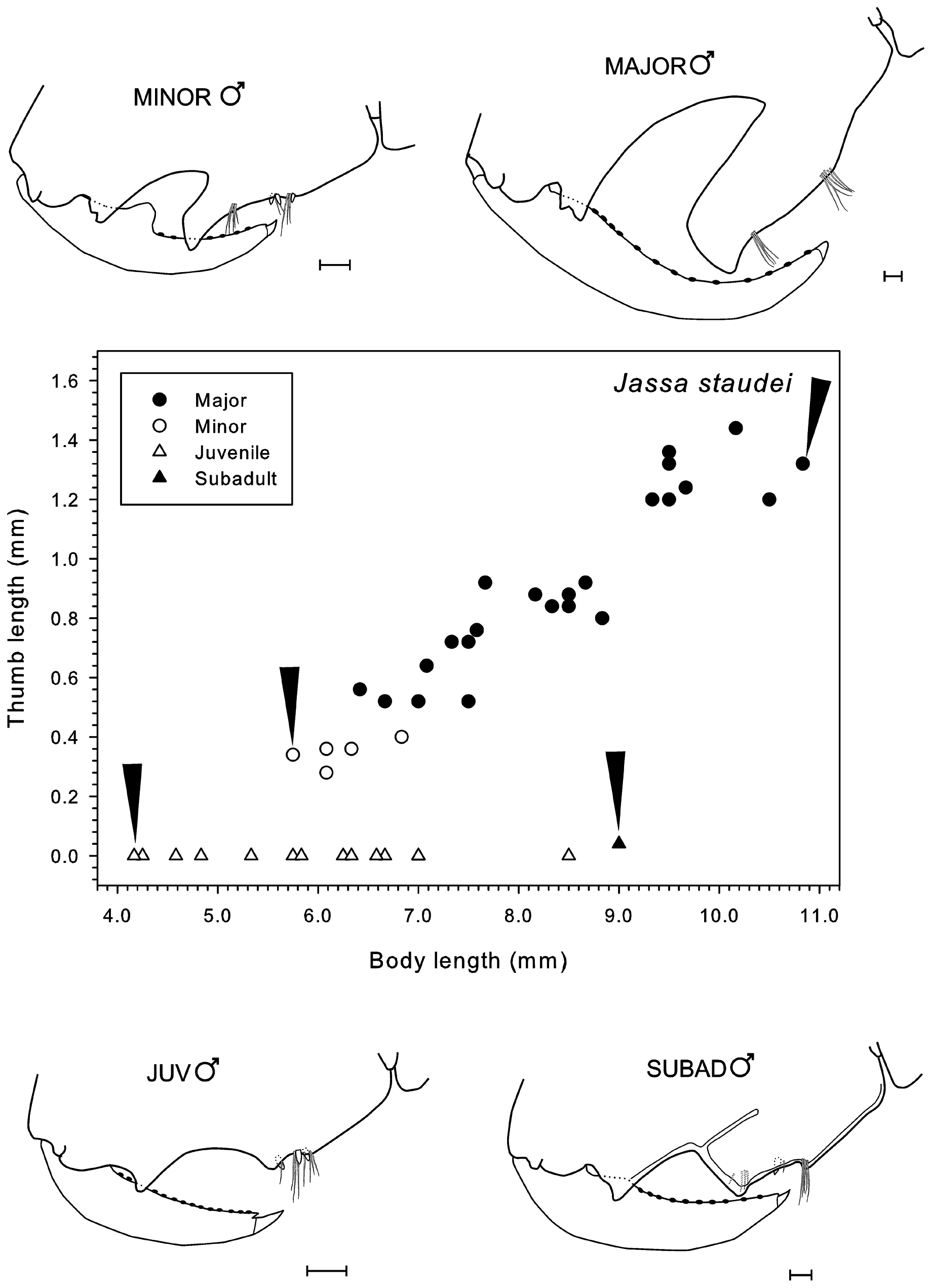
( Table 12 View TABLE 12 , Figs 67–77 View FIGURE 67 View FIGURE 68 View FIGURE 69 View FIGURE 70 View FIGURE 71 View FIGURE 72 View FIGURE 73 View FIGURE 74 View FIGURE 75 View FIGURE 76 View FIGURE 77 )
Diagnosis.
Both sexes:
Mandibular palp: article 2, dorsal margin without a fringe of setae.
Maxilla 1: without a seta or setal cluster at the base of the palp article 1.
Gnathopod 1: basis, anterolateral margin without a row of short setae along its length; carpus without a single or small cluster of setae at the anterodistal junction of the propodus.
Gnathopod 2: basis with a row of long setae along the anterolateral margin (setae length 100% of basis width); carpus and propodus, setae on the anterior margin long and simple (some setal lengths Ẑbasis width).
Pereopods 5–7: propodus not expanded anteriorly.
Uropod 1: ventral peduncular spinous process underlying 35-40% of the longest ramus.
Uropod 3: inner ramus without spines mid-dorsally (with only the single apical spine).
Telson: tip without apical setae (only the usual short setae at each dorsolateral cusp).
Thumbed male:
Antenna 2: large individuals with plumose setae on the flagellum and peduncular article 5 and lacking long filter setae.
Gnathopod 2: propodus, palmar defining spines not produced on a ledge, present in small thumbed males but absent in large thumbed males. In minor males, the thumb is distally squared, short relative to body length and located on the distal half of the propodus. The dactyl is strongly centrally toothed. In major males, the thumb is distally squared, longer relative to body length and on the proximal half of the propodus. The dactyl is expanded close to the junction with the propodus but is not centrally toothed.
Adult female:
Antenna 2: large animals with plumose setae on the flagellum and peduncular article 5.
Gnathopod 2: propodus, palm concave, palmar defining angle acute.
Remarks. Conlan (1990) noted that J. staudei has longer setae along the anteroproximal margin of the second gnathopod propodus than any other species. This is a key feature of the species as it is invariate (occurring in both sexes and at all ages). Another key feature is the lack of a single or cluster of setae at the junction of the carpus and propodus of gnathopod 1 (also an invariate character state). Other characteristics of the species are the plumose setae that appear on the antenna 2 peduncle article 5 and flagellum in larger males and females, the long, dense setae on the anterior margin of the gnathopod 2 basis, the well-developed peduncular spinous process on uropod 1, and the characteristics of the maxilla 1, mandibular palp and uropod 1 as noted above. All these characters combined will serve to distinguish this speci es from congeners on the Pacific coast of North America, whether native or introduced.
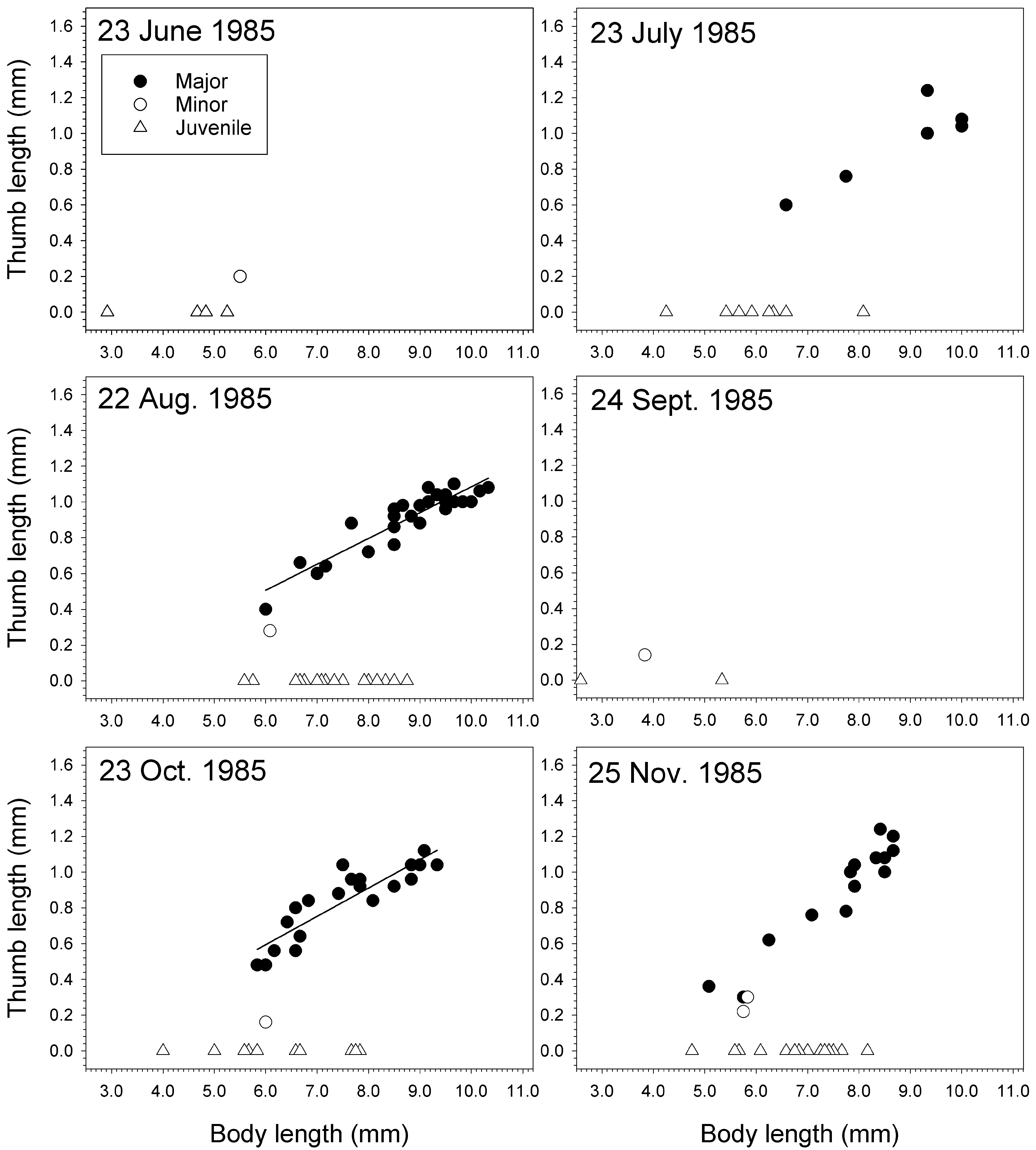
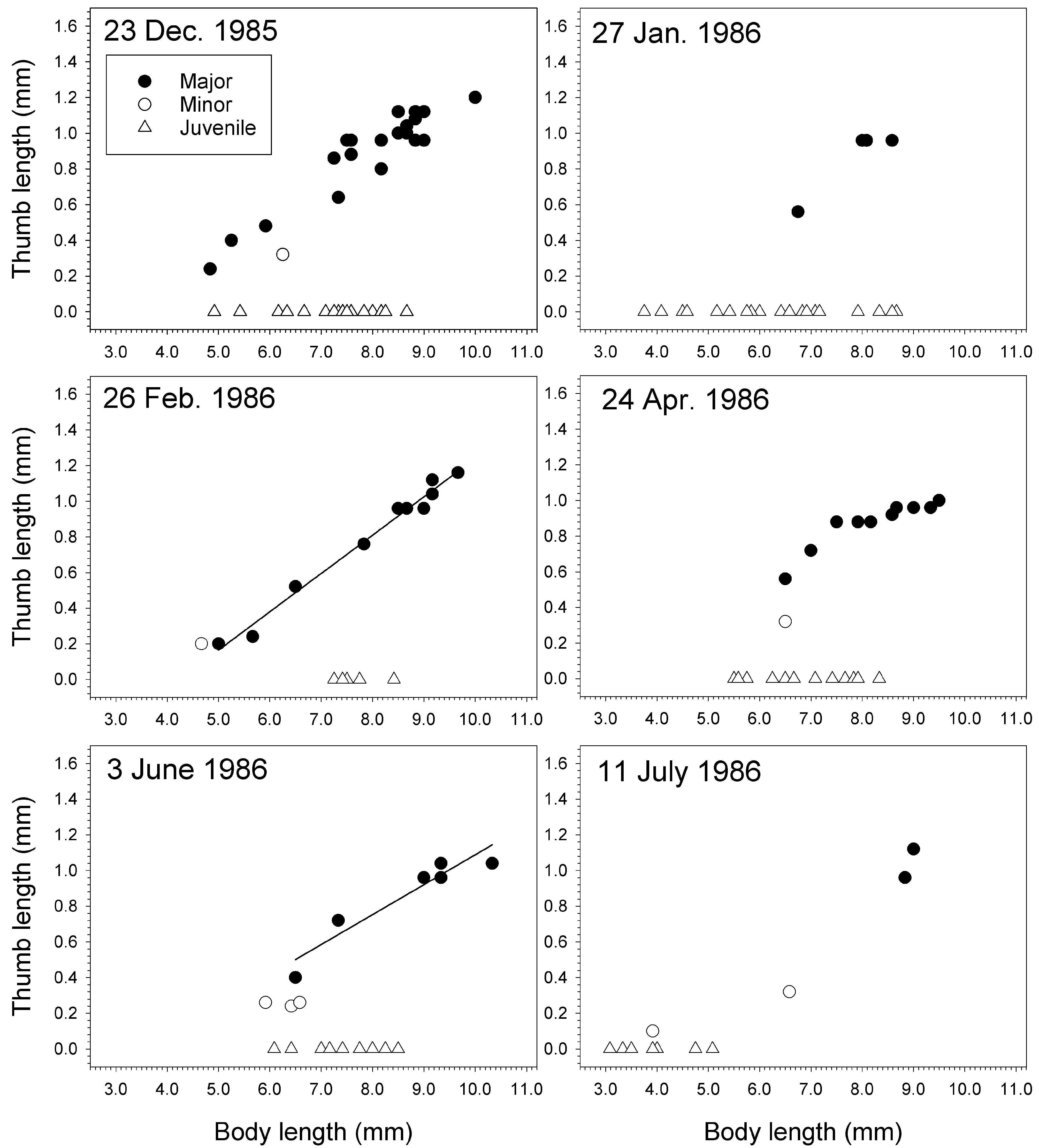
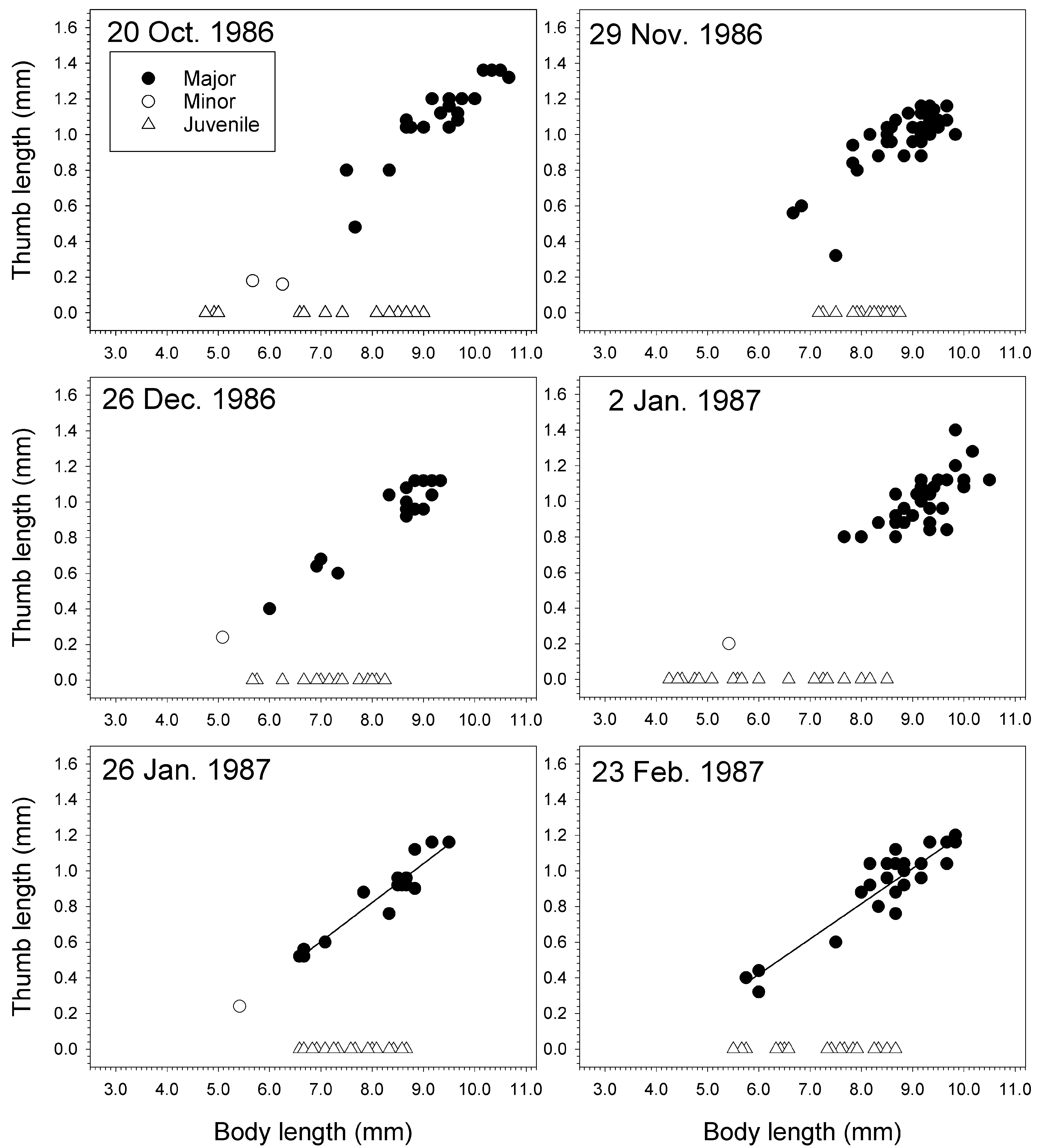
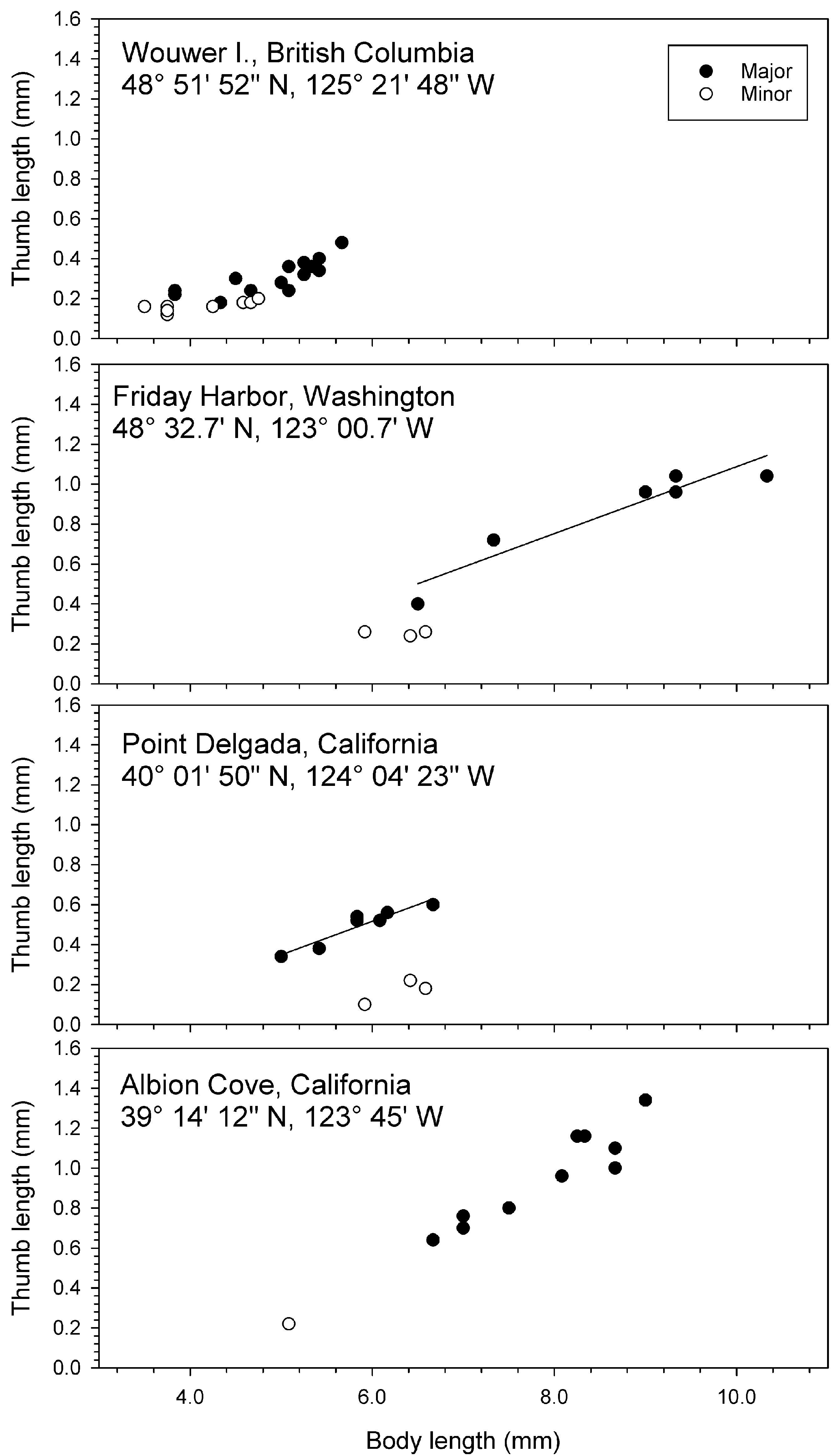
In a population of J. staudei collected inside the pressure head tank for the aquaria at Friday Harbor Laboratories, Washington, minor forms were rare compared to major forms ( Table 14 View TABLE 14 , Figs 69–72 View FIGURE 69 View FIGURE 70 View FIGURE 71 View FIGURE 72 ). Sampled near-monthly from June 1985 to Feb. 1987 ( Figs 70–72 View FIGURE 70 View FIGURE 71 View FIGURE 72 ), major forms achieved a greater body length than minor forms on average and always had longer thumbs relative to body length than the minor forms ( Table 14 View TABLE 14 ). Average major form body length was 8.5 ± 1.1 mm (n = 276), witha range of 4.8–10.7 mm. The smallest major form was found in December 1985 (4.8 mm) and largest in October 1986 (10.7 mm). Only 19 minor forms were found over the 18 samplings, giving an overall ratio for minor to major of 0.069:1. Average minor form body length was 5.7 ± 0.8 mm. The smallest minor form was found in September 1985 (3.8 mm) and the largest in June and July 1986 (6.6 mm). Juvenile males were found in all samplings with a maximum body length of 9.0 mm found in October 1986. However, large juveniles close to this size were found in most of the other collections as well .
Minor forms were also rare in four geographically varying populations collected in June along the western coast of North America ( Fig. 73 View FIGURE 73 ). Average body length of major form males was greatest at the protected location (Friday Harbor, Washington: 8.69 ± 1.31 mm). Among the three other locations, which were on exposed Pacific coasts, average body length increased southwards, from Wouwer Island , British Columbia (4.93 ± 0.57 mm) to Pt. Delgada, California (5.86 ± 0.50 mm) to Albion Cove, California (7.92 ± 0.82 mm) .
The Albion Cove population was also assessed for length of the gnathopod 2 propodus relative to body length ( Fig. 74 View FIGURE 74 ). For the four groups that were sufficiently numerous to measure, the relationship of propodus to body length appeared to be isometric for females and allometric for males. However, the data failed one or more assumptions of normality, constant variance or independence of residuals and so regressions were not applied.Adult major form males (n = 10) were considerably greater in body length (7.92 ± 0.82 mm, n = 10), than adult females (6.07 ± 0.79 mm, n = 19) and had a longer propodus as well (2.85 ± 0.45 mm vs 1.28 ± 0.19 mm). Of the 23 juvenile males measured, only one overlapped the major form adult males in body length (7.17 mm) but its propodus length (1.84 mm) was shorter than any of the major form adults that were of similar size (propodus length 2.31 ± 0.12 mm, body length 6.89 ± 0.19 mm, n = 3). For the females, there was no overlap in body length between juveniles and adults. However, for juvenile females and juvenile males, propodus length was similar among small sized individuals of both sexes but diverged in larger juveniles with a longer propodus in the males than females. However, the differences in the median propodus length between the juvenile males (n = 8) and adult females (n = 19) that overlapped in body length (5.0–8.0 mm) were not great enough to exclude the possibility that the difference was due to random sampling variability (ANOVA, F = 3.651, p = 0.068, power = 0.332).
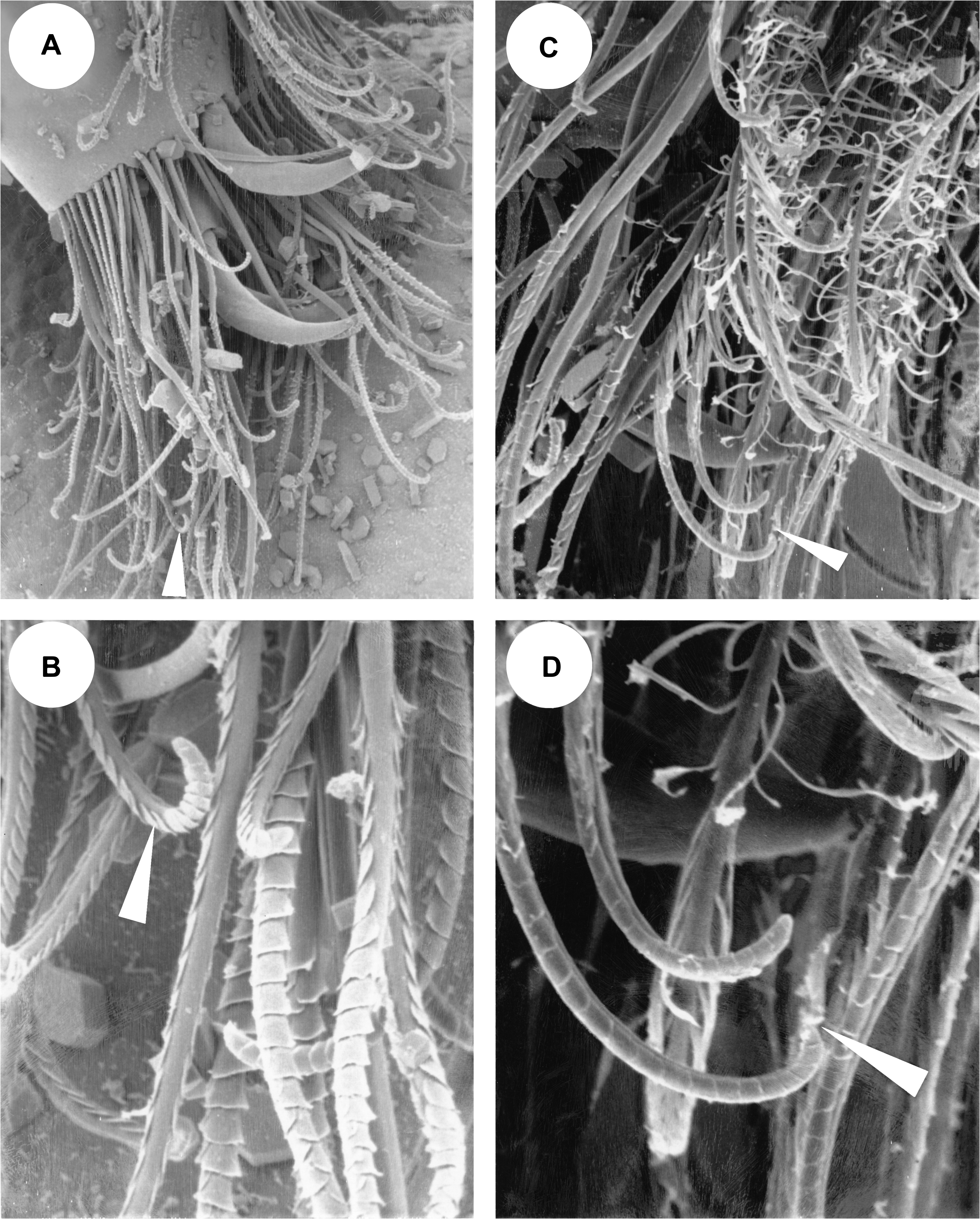
Fig. 76 View FIGURE 76 shows magnifications of the second antenna flagellum in an adult female (A, B) and a subadult male (C, D). For suspension feeding, the second antennae are splayed out ~90°to either side of the body and the antennal setae spread widely. Along with the splayed first antennae, this provides a near 180°wide net for the capture of drifting particles. The curved spines at the flagellum tip (A) and scaled setae (B) are adaptations for deposited (spines) and suspended (scales) food capture. Fig. 76B View FIGURE 76 shows that the scales cover the anterior length of the filter setae and wrap around the setae only partially, so the posterior length is bare. Possibly this arrangement improves capture of small particles which are then scraped by the medial setae of the first and second gnathopods, mandibular palps and maxilliped palps. The antennal setae are also used to capture particles on the uropods, when the urosome is bent under the body toward the head (pers. obs. on J. marmorata ). The plumose setae typical of larger individuals of both sexes are visible in the subadult male ( Fig. 76C, D View FIGURE 76 ). The tip of the plumose seta, visible in Fig. 76D View FIGURE 76 , has unextended plumes, giving the tip a bulbous appearance. The function of these plumose setae is presumably for sexual sensing since it occurs in adults and subadults but not juveniles. Plumosity can be dense in some species (e.g., in J. staudei , J. falcata , J. slatteryi , J. oclairi , J. borowskyae ), though in all species there is a modification of setation on the posterior part of the antenna 2 flagellum at least in the adult males.
Fig. 77 View FIGURE 77 shows an unusual cusping at the tip of the third uropod outer ramus with four cusps rather than the usual two proximal to the basally immersed, dorsally recurved spine.
Piola & Conwell (2010) (repeated by Marchini & Cardeccia 2017) reported J. staudei fouling a fishing boat docked in New Zealand (location not given). It had been identified by NIWA’s Marine Invasives Taxonomic Service (MITS) but this identification has not been confirmed. If it is correct, this would be the first instance of J. staudei having been found outside the North Pacific. The point of origin of the fishing boat was not indicated. The study embraced fishing boats only from Australia / New Zealand, the Northwest Pacific, and the South Pacific. If the fouling species was J. staudei , it would have been likely that the fishing boat was from J. staudei ’s home region, the Northwest Pacific. However, the two boats sampled from this region recorded no fouling. Jassa staudei is not known from either of the other two regions. It is possible that the specimen(s) were mis-identified and were more likely J. marmorata or J. slatteryi , which have been confirmed present in New Zealand since 1928 (Table 3). Jassa marmorata was found fouling two other fishing boats in the study. Marchini & Cardeccia (2017) cited Inglis et al. (2008) as finding J. staudei in the Port of Timaru, New Zealand. Inglis et al. (2008) were referring to J. slatteryi , however; no mention of J. staudei could be found.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.