
Parethusa hylophora, Castro, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.5399909 |
|
persistent identifier |
https://treatment.plazi.org/id/264A053E-4E75-B57A-71E3-FCC776D3C37C |
|
treatment provided by |
Marcus |
|
scientific name |
Parethusa hylophora |
| status |
sp. nov. |
Parethusa hylophora View in CoL n. sp.
(Fig. 32)
T Y P E M A T E R I A L. — Holotype: cl 5.2 mm, c w 4. 7 m m, S A L O M O N 1, s t n C P 1 8 5 1 ( MNHN-B 28667 ); paratype: cl 5.1 mm, cw 4.7 mm, SALOMON 1, stn CP 1783 ( MNHN-B 28716 ).
MATERIAL EXAMINED. — Solomon Islands. SALOMON 1, stn CP 1783, 08°32.8’S, 160°41.7’E, 399-700 m, 29.IX.2001, 1 paratype ( MNHN-B 28716). — Stn CP 1851, 10°27.6’S, 162°00’E, 297-350 m, 6.X.2001, 1 holotype ( MNHN-B 28667).
TYPE LOCALITY. — Solomon Islands, N of San Cristobal island, 10°27.6’S, 162°00’E, 297- 350 m.
ETYMOLOGY. — From hyle (Greek for wood) and - phor (Greek suffix meaning to carry) in reference to the holotype being collected under a piece of wood.
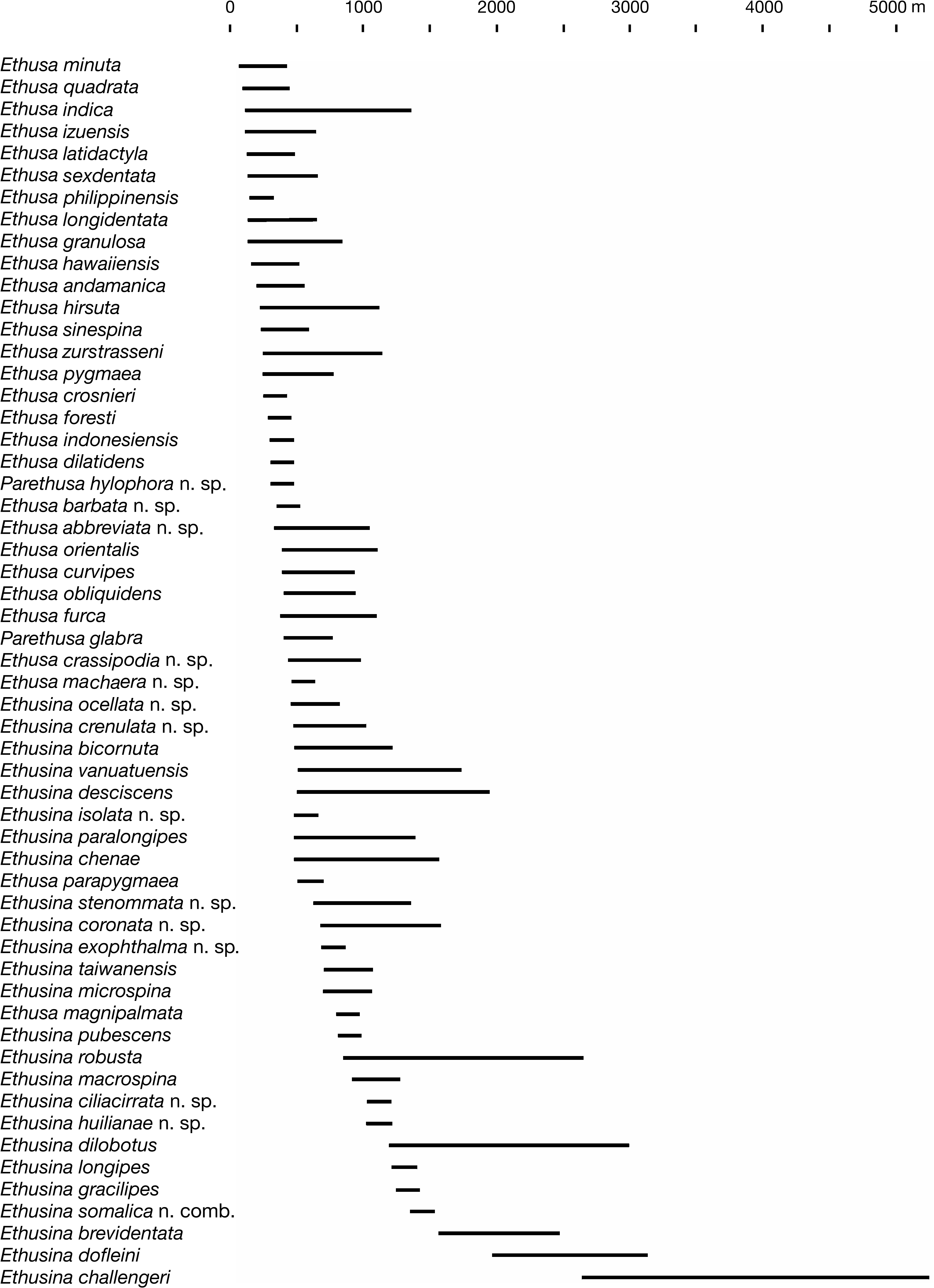
DISTRIBUTION. — Known only from the Solomon Islands. Depth: 297-700 m ( Fig. 34 View FIG ).
SIZE. — Maximum size: 5.2 cl mm, cw 4.7 mm (MNHN-B 28667), unknown.
DESCRIPTION
Carapace (Fig. 32A) slightly longer than broad in males (females unknown); dorsal surface smooth, flat; carapace regions indistinct, only urogastric region defined by very slight groove. Nearly straight lateral borders that diverge toward posterior border, slight indentation at the level of branchial groove.
Anterior border of carapace (Fig. 32A) with four teeth. Outer orbital teeth triangular, straight, longer but not reaching frontal teeth. Lateral frontal teeth with oblique anterior borders almost parallel to anterior edge of carapace, much short- er than median frontal, outer orbital teeth. Median frontal teeth oblique, obtuse tip. Orbital sinuses V-shaped; lateral frontal sinuses oblique, U-shaped; median frontal sinus V-shaped, wider than lateral frontal and orbital sinuses.
Eyes ventral, visible dorsally (Fig. 32B). Eye peduncles twice eye diameter, mobile, visible dorsally.
Basal antennular article relatively large and swollen. Anterior border of endostome (Fig. 32B) lies well below posterior border of antennular fossae of basal antennular articles.
Male chelipeds (P1) (females unknown) smooth, equal; slender propodi, slightly longer than fingers, with broad cutting edges.
P2, P3 relatively short; length of P2 meri 0.9 times cl, P2 meri 8.0 times longer than broad. P4, P5 with few short hairs; P5 (Fig. 32C) dactyli thick, very strongly curved.
Male abdomen (females unknown) with four somites (3-5 fused, basal half swollen), rounded telson. Somite 1 length 2.1 times as broad, somite 6 square.
G1 (Fig. 32D) short, stout; each distal end with slight dorsolateral swelling with gonopore, point- ed tip fringed with spines; G2 (Fig. 32E) relatively short, slightly sinuous, each distal parts narrower; pointed tip.
REMARKS
Parethusa hylophora View in CoL n. sp. is distinguished from P. glabra Chen, 1997 View in CoL , the other known species in the genus, by the presence of four frontal teeth instead of the two in P. glabra View in CoL ( Chen 1997: fig. 1a, b). Only two specimens of the new species are known, both males. The lateral borders of the carapace of the males of P. hylophora View in CoL n. sp. are straight, and diverge toward the posterior border in contrast to the inflated lateral borders of male and female P. glabra View in CoL ( Chen 1997: fig. 1a). The
Fig. 32. — Parethusa hylophora View in CoL n. sp., holotype cl 5.2 mm, cw 4.7 mm, Solomon Islands, north of San Cristobal, SALOMON 1, stn CP 1851, 297- 350 m ( MNHN-B 28667 ) ; A, dorsal surface of carapace; B, ventral view of anterior portion of body; C, P5, dorsal view; D, left G1, ventral views (left and right), dorsal view (center); E, left G2, ventral view. Scale bars: A, 5.0 mm; B, E, 1.0 mm; C, 2.0 mm; D, 0.5 mm.
eyes of P. hylophora n. sp. lack the thick, ventral process characteristic of the eyes of P. glabra ( Chen 1997: fig. 1b).
The holotype (MNHN-B 28667) was found under a piece of wood that had been dredged north of San Cristobal island in the Solomon Is. In this location, trees are easily carried into deep water along coasts that are not bordered by the shallow lagoons of fringing reefs (P. Bouchet pers. comm.).
CARRYING BEHAVIOUR AMONG INDO-WEST PACIFIC ETHUSINES
There is very little information on the ability of ethusines to carry an object with the hook-like dactyli of their P4 and P5. Crabs most probably release any objects they may have been carrying during collection so there are very few reliable records. Some of the few observations that do exist are from crabs kept in aquaria after collection. The closely-related, shallow-water dorippid crabs (subfamily Dorippinae ), however, are known to carry a large variety of organisms, even leaves, with their dorsal, highly mobile, subchelate P4 and P5 ( Fenizia 1937; Wicksten 1986; Holthuis & Manning 1990; Guinot et al. 1995; Chen & Sun 2002). They are commonly known as “carrier” or “porter” crabs.
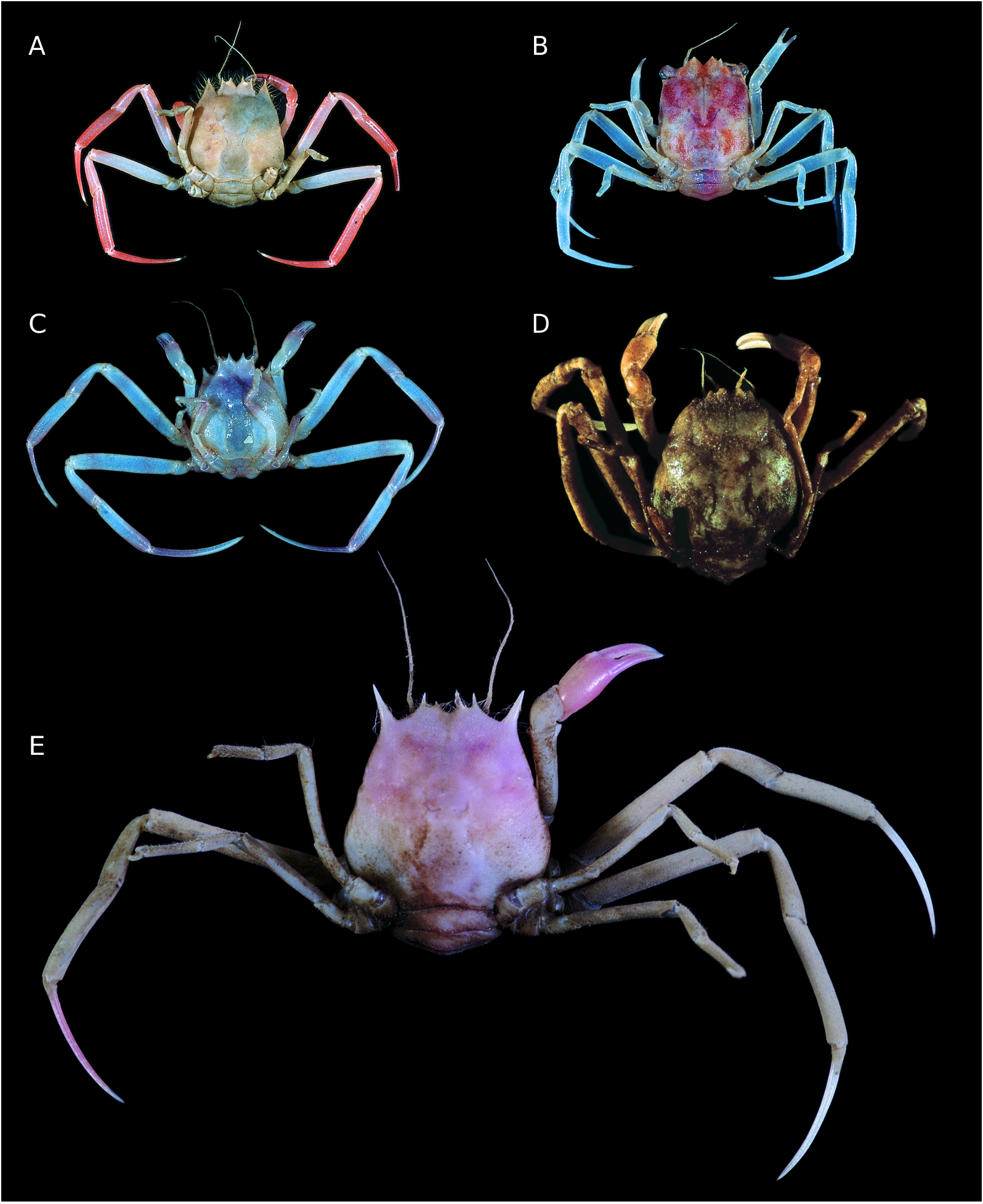
Photos show an individual of Ethusa mascarone (Herbst, 1785) , a Mediterranean and Atlantic species, covering itself with a bivalve shell ( Gruner 1969). Sakai (1937: 80) commented on Ethusa izuensis Sakai, 1937 using bivalve shells in Japan. Individuals of Ethusa sexdentata ( Stimpson, 1858) kept in an aquarium in Japan were also observed taking and carrying a bivalve shell or small pieces of wood with their P4 and P5 (T. Komai pers. comm.). One of the two known specimens of Parethusa hylophora n. sp. was collected under a piece of wood in the Solomon Is.
There are no obvious differences in the morphology of the P4 and P5 among ethusines that would suggest morphological adaptations for carrying particular objects. Differences in carapace size observed among species, however, most probably call for for the carrying of different sizes and types of objects.
DEPTH DISTRIBUTION OF INDO-WEST PACIFIC ETHUSINES
There are striking differences between the three genera of ethusines in terms of their depth distributions. Practically all species of Ethusa and Parethusa have been collected from depths that are lower than the shallowest values ever recorded for Ethusina species ( Fig. 34 View FIG ). Only two of the 30 species of Ethusa ( E. hirsuta and E. indica ) and Parethusa have been found in material collected at depth of more than 1000 m. Conversely, only five of the 25 species of Ethusina ( E. exophthalma n. sp., E. ocellata n. sp., E pubescens , E. vanuatuensis , and E. crenulata n. sp.) have never been found in samples collected at depths greater than 1000 m. This may certainly change with more extensive collecting.
Miers (1886: 332), in his description of E. challengeri , commented that at 1875 fathoms (3429 m), the holotype was obtained from “the greatest depth hitherto recorded for any species of Crab”. A female specimen of the same species was dredged from 4928-4950 m in the Indian Ocean (MNHN-B 28697; Fig. 33D View FIG ). Two specimens of Ethusina abyssicola Smith, 1884 , however, were collected from a depth of 5046 m in the Caribbean Sea ( Gore 1983: 213). Both E. challengeri and E. abyssicola are therefore true abyssal species, having been found at depths exceeding 4000 m.
BIOGEOGRAPHY OF INDO-WEST PACIFIC ETHUSINES
Little can be said about the biogeography of Indo-West Pacific ethusines because of the relatively limited distribution of deep-water collections. Nevertheless, some patterns emerge from what we know, particularly in the Western Pacific, where collections have been relatively more frequent and extensive than elsewhere in the region.
Most species of ethusines show a relatively narrow geographical distribution. It may be speculated that such pattern may result from short larval development. The most obvious exception of a limited distribution is Ethusa indica , which has been collected from the Gulf of Aden (and possibly the Red Sea) to the Western Pacific from Japan to Fiji ( Fig. 12 View FIG ), a distributional pattern followed by many Indo-West Pacific shallowwater brachyurans. Ethusina challengeri also shows a wide although discontinuous distribution in both oceans ( Fig. 17 View FIG ), most probably because it is rarely collected due to the great depths it inhabits ( Fig. 34 View FIG ).
Very few other ethusines are known from both the Indian and Pacific oceans: Ethusa hirsuta ( Fig. 11 View FIG ), Ethusina dofleini , Ethusina gracilipes , and Ethusina robusta ( Fig. 22 View FIG ).
What may appear as peripheral endemicity along the southwestern and western Indian Ocean (for Ethusa machaera n. sp., E. sinespina , E. zurstrasseni , Ethusina longipes , and E. somalica ) and the eastern limits of the Indo-West Pacific ( Ethusina crenulata n. sp., E. huilianae n. sp., and E. isolata n. sp.) could be the result of the more intense collecting in these areas in contrast to very few from most of the Indian Ocean and the central Pacific, respectively.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parethusa hylophora
| Castro, Peter 2005 |
Parethusa hylophora
| Castro 2005 |
P. hylophora
| Castro 2005 |
Parethusa hylophora
| Castro 2005 |
P. glabra
| Chen 1997 |
P. glabra
| Chen 1997 |
P. glabra
| Chen 1997 |