
Temnocephala pignalberiae Dioni, 1967
|
publication ID |
https://doi.org/ 10.5281/zenodo.197863 |
|
DOI |
https://doi.org/10.5281/zenodo.6203218 |
|
persistent identifier |
https://treatment.plazi.org/id/03E6C930-FFAF-FFF5-09BF-FF1DFB5EFAD9 |
|
treatment provided by |
Plazi |
|
scientific name |
Temnocephala pignalberiae Dioni, 1967 |
| status |
|
Temnocephala pignalberiae Dioni, 1967 View in CoL
(Figs. 3–37)
Study based on 57 specimens collected: 26 specimens stained in hematoxylin mounted in toto: 8 from Poconé and 18 from Bebedouro; 6 specimens stained in aceto-carmine/fast green mounted in toto: 5 from Poconé and 1 from Bebedouro; 22 extracted cirri mounted in Faure´s medium: 5 from Poconé and 17 from Bebedouro; and 3 slides with eggs mounted in Canada balsam: 2 from Poconé and 1 from Bebedouro.
Table 1 presents morphometric data comparing specimens measured from Poconé and Bebedouro.
Taxonomic summary. Type host and type locality: Dioni (1967) did not indicate the type host and the type locality for this species: “ Trichodactylus orbicularis Meuschen , Río Salado, Santo Tomé, Argentina; Trichodactylus pictus, Madrejón Don Felipe y Laguna Guadalupe, Santa Fe, Argentina; Trichodactylus sp., Madrejón Don Felipe, Santa Fe; Trichodactylus sp., alrededores de Paraná, Entre Ríos; T. pictus y T. orbicularis, Madrejón Don Felipe , Santa Fe; T. pictus, Laguna Guadalupe, Santa Fe”.
Other hosts and localities: Argentina: “ Dilocarcinus pagei Stimpson, 1861 , Laguna Guadalupe (Piedras Blancas), Santa Fe, Argentina ( Damborenea, 1992)”. Brazil (present work): Dilocarcinus pagei in Poconé, State of Mato Grosso and in Bebedouro, State of São Paulo.
Site of infestation: adults found inside the branchial chambers; eggs deposited over the tissue covering the inner upper part of the carapace.
Prevalence: 100%, in both localities, Poconé, MT and Bebedouro, SP.
Specimens deposited: CHIOC Nº 37307 (SBA- 2801-2-2, eggs), CHIOC Nº 37308 (SBA- 2807-1-3, specimen in toto), CHIOC Nº 37309 (SBA- 2799-1-5, cirrus in F), CHIOC Nº 37310 a-b (SBA-3076-1-1 (a), specimen in toto; SBA-3076-1-6 (b), specimen in toto), CHIOC Nº 37311 (SBA- 2944-1-1, cirrus in F); INPA Nº 525 (SBA-3076-1-5: specimen in toto), INPA Nº 526 (SBA- 2945-1-4: cirrus in F), INPA Nº 527 (SBA- 2799-1-3: specimen in toto), INPA Nº 528 (SBA- 2800-1-2: cirrus in F); MLP Nº 6091 (SBA- 2807-1-1: specimen in toto), MLP Nº 6092 (SBA- 2799-1-4, cirrus in F), MLP Nº 6093 (SBA- 2802-2-1, eggs), MLP Nº 6094 (SBA- 2946-1-4: specimen in toto), MLP Nº 6095 (SBA- 2945-1-3, cirrus in F).
Specimens examined: “ Temnocephala pignalberiae , MLP Nº 3126, camaras branquiales, Dilocarcinus (D.) pagei, Laguna Guadalupe, Santa Fe, Argentina, col. 2 Febrero 1987 ”; “ Temnocephala pignalberiae , MLP Nº 3127, camaras branquiales, Sylviocarcinus pictus, Laguna Guadalupe, Santa Fe, Argentina. Col. 2 Febrero 1987 ”; “ Temnocephala pignalberiae , MLP Nº 3127, camaras branquiales, Sylviocarcinus pictus, Laguna Guadalupe (Piedras Blancas), Santa Fe, Argentina. Col. 1 Febrero 1987 ”; and “ Temnocephala pignalberiae , MLP” (slide without collection number, host name, date of collection, and collector indication).
Host specimens deposited: “Coleção de Crustáceos UFRGS Nº 4565 – SBA 3184 – Dilocarcinus pagei, Bebedouro Col. R. J. Ilário ”; “Coleção de Hospedeiros Laboratório de Helmintologia UFRGS – SBA 3185 – Dilocarcinus pagei, Bebedouro. Col. R. J. Ilário. ”
Comments. Dioni (1967) did not indicate having elected and deposited a holotype and paratypes of T. pignalberiae ; consequently, he also did not indicate which of the two species of crabs, D. pagei or S. australis , would be the type host, nor did he indicate which of the five collection localities would be the type locality for the species. Later, Damborenea (1992) and Damborenea & Cannon (2001) collected again T. pignalberiae from the same hosts and from the same localities in Argentina.
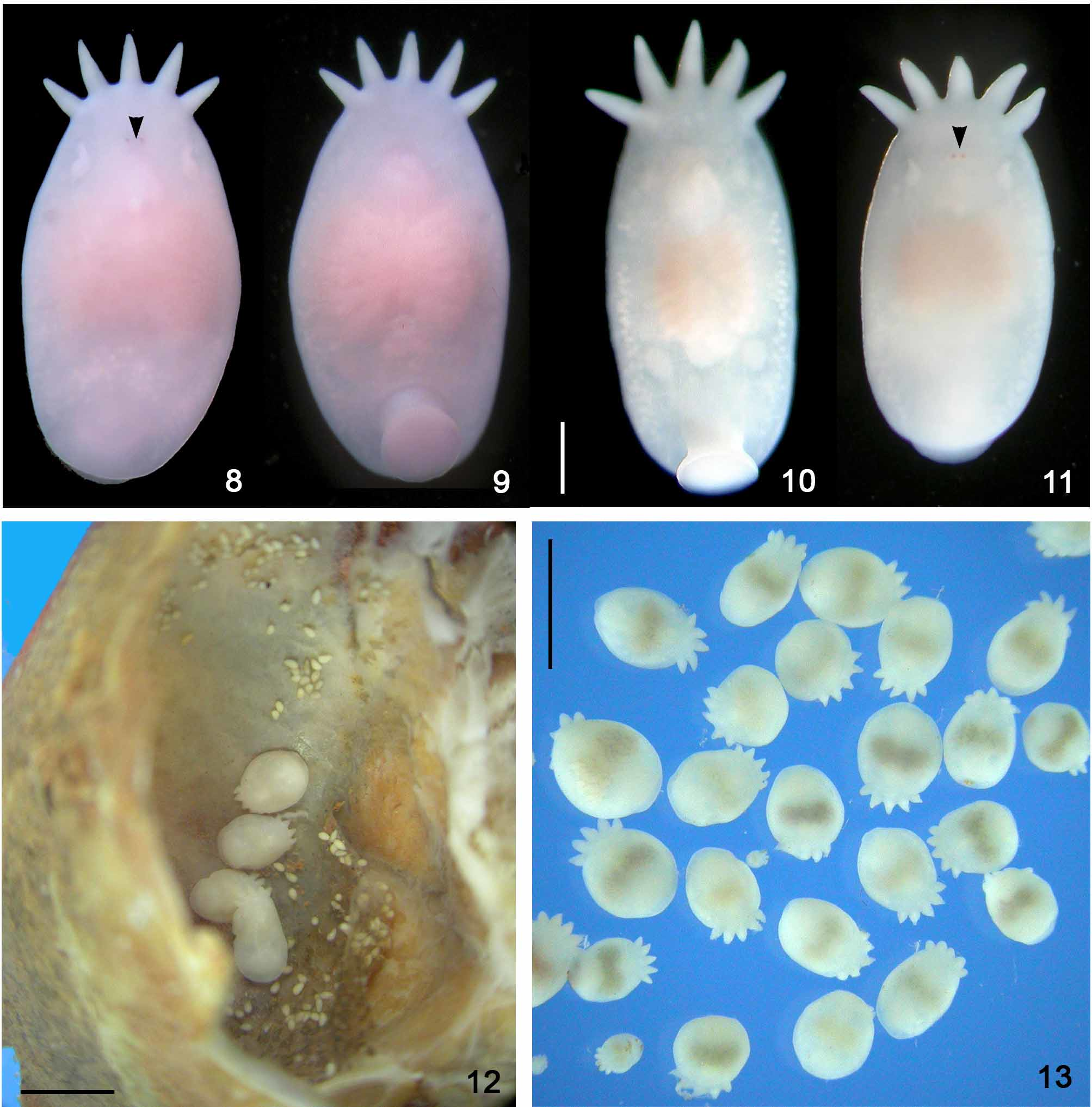
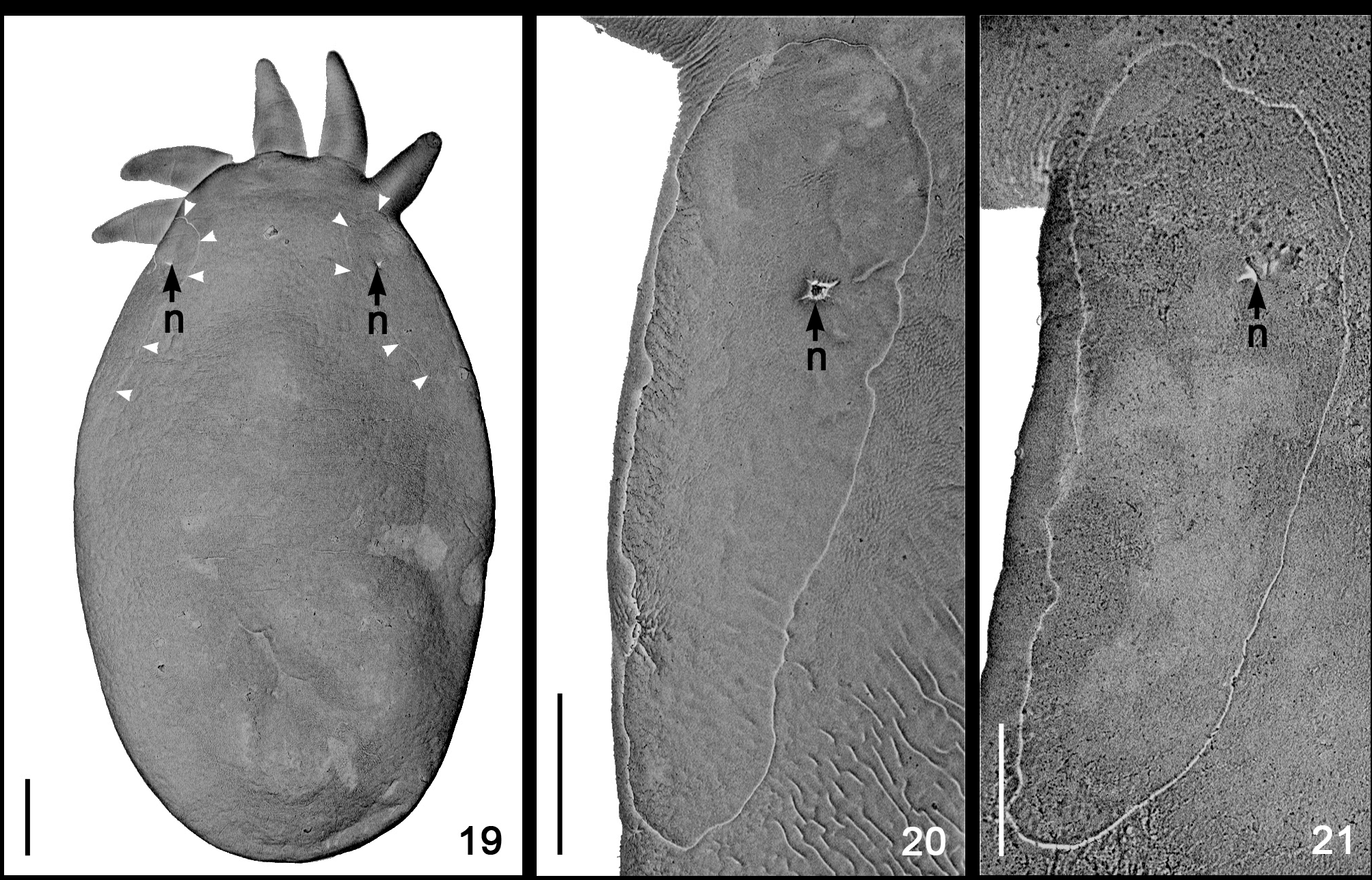
Temnocephala pignalberiae is the species with the most perfect elliptical body shape (when properly killed/fixed by formalin 10% formalin 90ºC — compare Figs. 8–11 View FIGURES 8 – 13 with Figs. 12 and 13 View FIGURES 8 – 13 ) and the smallest tentacles, among the species so far studied. The crabs infested with T. pignalberiae did not have temnocephalan eggs deposited on the external areas of the carapace, abdomen or pereiopods ( Figs. 1 and 2 View FIGURES 1 – 2 ). The eggs laid by T. pignalberiae are the most delicate which we have found in any of the species studied. Additionally, these eggs being laid over the tissue which covers the inner upper surface of the carapace were collected by ‘peeling off’ this tissue (Figs. 4 and 5). These eggs have a long, subterminal filament (Fig. 6). In the present paper, specimens of D. pagei , from Poconé and Bebedouro, were infested exclusively by T. pignalberiae , which allowed us to be certain about the morphology and the correct place for the deposition of eggs by this species.
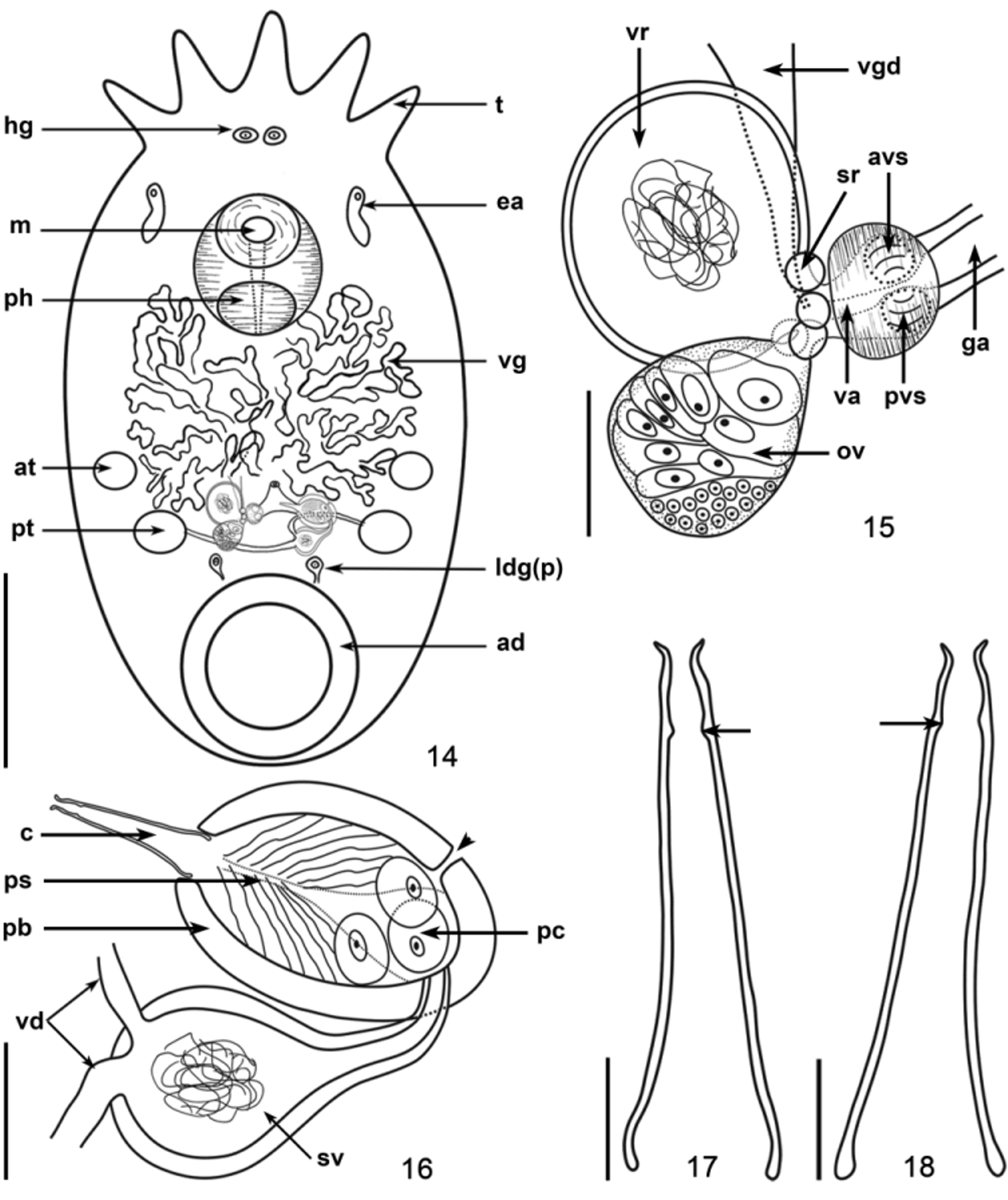
Specimens of T. pignalberiae from these two allopatric populations of D. pagei showed many similarities when compared: above all, the shape of the cirrus, which is the only unarmed (without spines in the introvert) cirrus found in species of Temnocephala . The cirri in specimens from Poconé are proportionally longer, in relation to the length of the prostatic bulbs. The latter have thicker walls and are less elongate than those found in specimens from Bebedouro. In specimens from Poconé the proximal end of the shaft tends to have the rims curved inward, when compared to the rims of the shaft of cirri in the specimens from Bebedouro. The prostatic bulb is more elongated (pyriform) than elliptical, and the rims of the proximal extremity of the shaft are not curved inward ( Figs. 17 and 18 View FIGURES 14 – 18 ; 24 and 29).
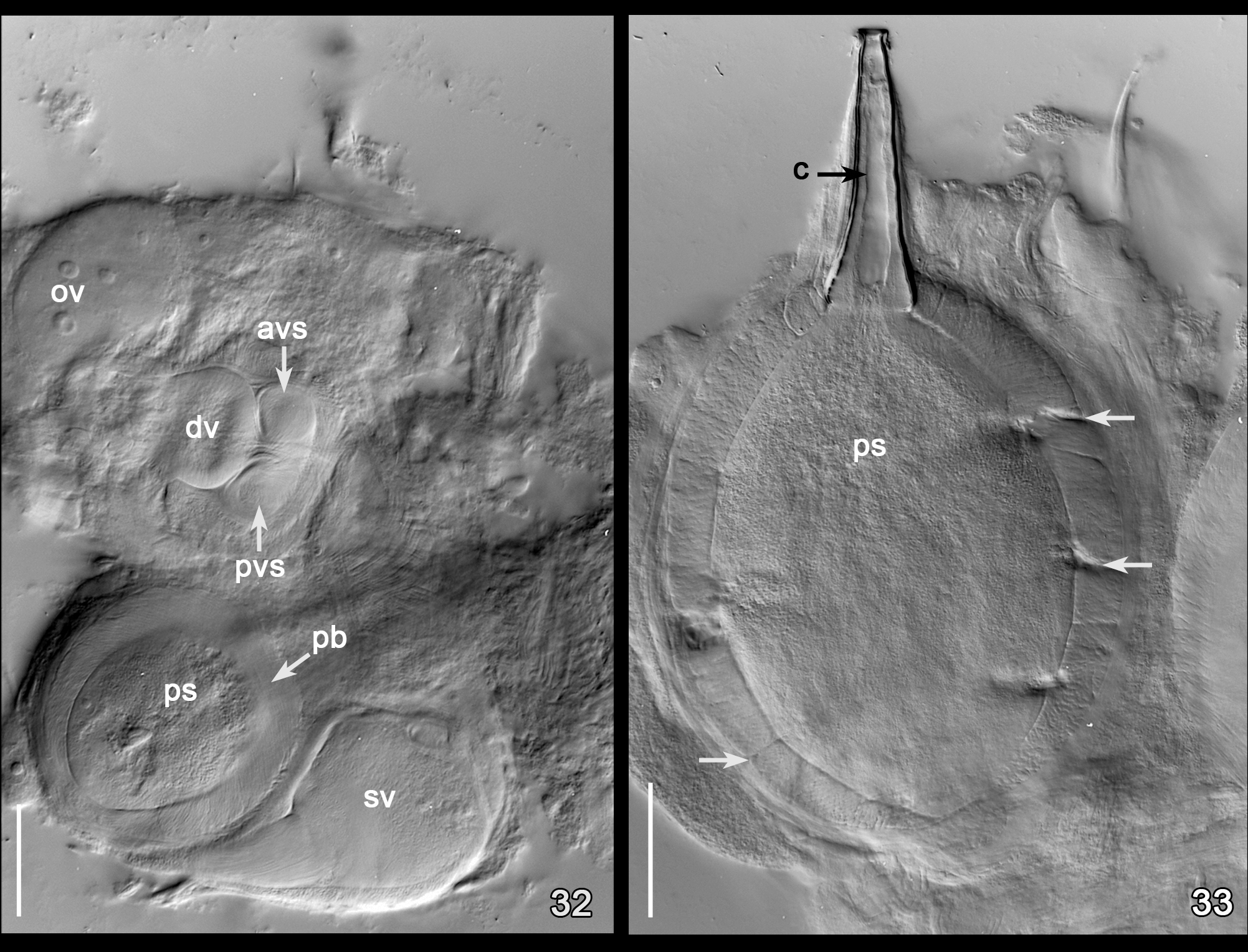
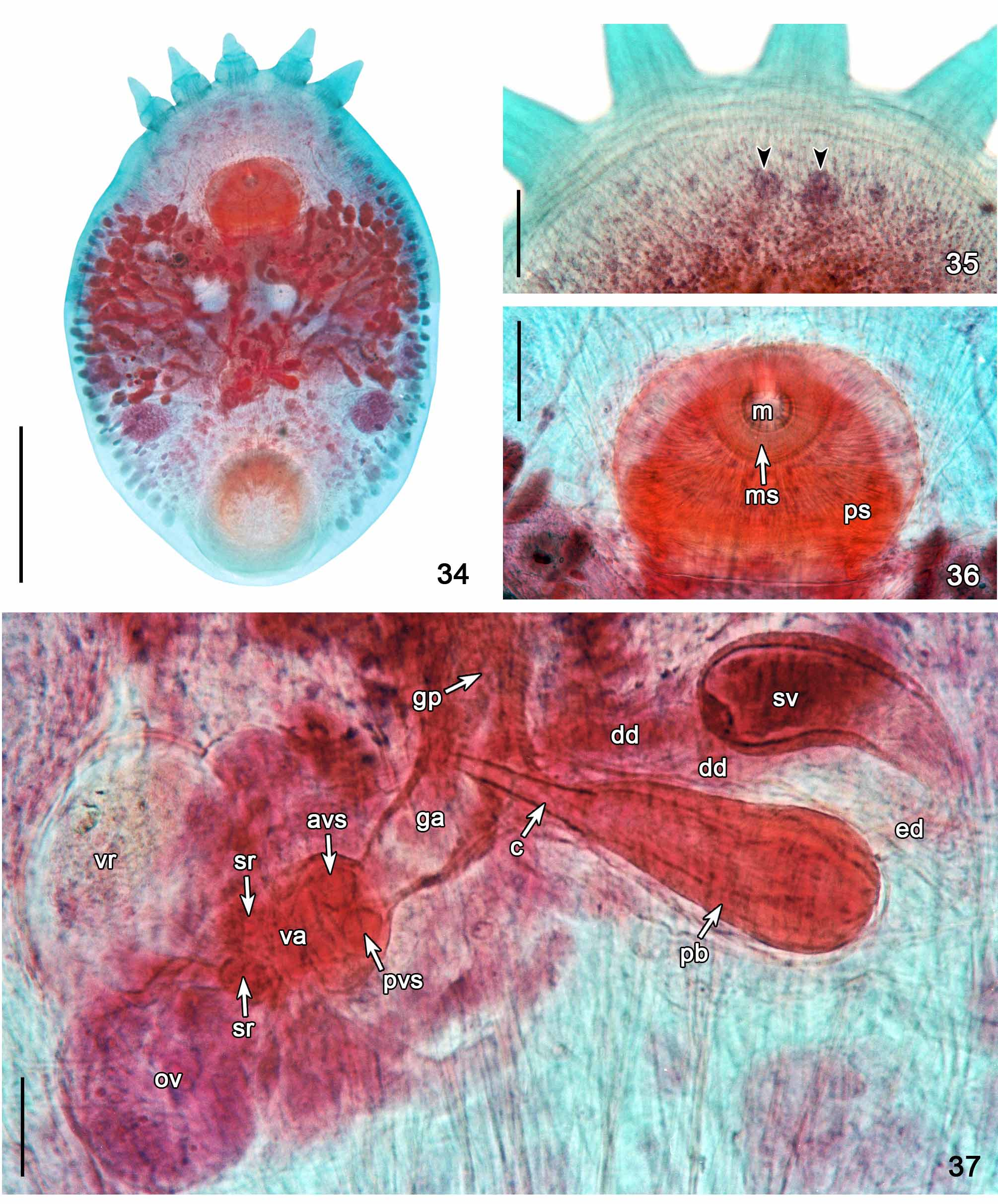
Temnocephala pignalberiae has only one vaginal sphincter, which appears to be double when observed in whole mounts, because the distal portion of the vagina has very thick walls, and can be clearly observed when DIC illumination is applied to dissected reproductive parts mounted in Faure´s medium ( Fig. 32 View FIGURES 32 – 33 ) or in specimens stained with aceto-carmine/fast green ( Fig. 37 View FIGURES 34 – 37 ).
Other similarities observed between the specimens from D. pagei , of Poconé and Bebedouro were the shape of the DLSPs, as revealed by SEM ( Figs. 20–21 View FIGURES 19 – 21 ); the presence of two large disk glands (paranephrocytes?); and the dorsal position of the vitellarium, which is well superposed to the intestinal sac and has the terminal portions of the branches turning toward its ventral side (Figs. 7 and 34).

| UFRGS |
Universidade Federale do Rio Grande do Sul |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.