
Dicerorhinus jeanvireti Guérin, 1972
|
publication ID |
https://doi.org/ 10.5252/g2013n2a7 |
|
persistent identifier |
https://treatment.plazi.org/id/D11D0148-5B0A-2F74-FD54-F32BFB06FE5D |
|
treatment provided by |
Marcus |
|
scientific name |
Dicerorhinus jeanvireti Guérin, 1972 |
| status |
|
Dicerorhinus jeanvireti Guérin, 1972
Dicerorhinus jeanvireti Guérin, 1972: 133-136 , pl. I; 1980: 607, 608. — Hooijer 1981: 472; 1985: 35-37. — Holec 1996: 219-222. — Durisova 2004: 99.
Rhinoceros elatus Croizet & Jobert, 1828: 144-154 . In pursuance of the article 23.12 of the International Code of Zoological Nomenclature, nomen oblitum because explicitly rejected by Guérin (1972), i.e. between 6 th November 1961 and 1 st January 1973, following the article 23b then in force between those dates.
Atelodus aymardi – Pomel 1851-52: 80, nomen nudum.
Rhinoceros mesotropus – Aymard in Pictet 1853: 298, nomen nudum.
Rhinoceros velaunus – Aymard in Pictet 1853: 298, nomen nudum.
Rhinoceros etruscus – Falconer 1868: 367. — Depéret et al. 1923: 37.
Rhinoceros etruscus var. astensis – Sacco 1895: 1-31, pls I-IV. Owing to the article 23.12 of the International Code of zoological nomenclature, nomen oblitum because explicitly rejected by Guérin (1972), i.e. between 6 th November 1961 and 1 st January 1973, following the article 23b then in force between those dates.
Rhinocéros de Vialette – Viret 1954: 154, text-fig. 35 and 42.
Dicerorhinus cf. megarhinus – enius 1955: 10, 11, pl. I, figs 1-4 (non Rhinoceros megarhinus de Christol, 1834 ).
Rhinoceros aff. megarhinus – Bout 1960: 188, 189.
Rhinoceros sp. – Bout 1960: 71.
Dicerorhinus megarhinus – Kurtèn 1963: 10, table 1. — Fejfar 1964: 89-101, fig. 47-55.
Rhinoceros megarhinus – Azzaroli 1963: 17.
“À Vialette, forme qui ressemble plutôt au Rhinoceros megarhinus de Montpellier qu’au Rhinoceros etruscus ” – Hürzeler 1967: 635.
Rhinoceros megarhinus astensis – Hürzeler 1967: 636.
Stephanorhinus jeanvireti – Campanino et al. 1994: 452, 453. — Mazo 1995: 287. — Gliozzi et al. 1997: 171, table 1.l. — Spassov 2005: 203, 204. — Lacombat & Mörs 2008: 160.
Stephanorhinus cf. elatus in the sites of Covrigi, Iaras 2 and Ilieni – Radulescu et al. 2003: 3, 8.
S. elatus in the sites of Groserea, Cernutesti and Araci- Fântâna Fagului. — Radulescu et al. 2003: 3, 4, 8.
Stephanorhinus elatus – Radulescu 2005: 193. THE RUSCINIAN AND VILLAFRANCHIAN EUROPEAN RHINOS
Four species of rhinoceros are known in the Ruscinian and Villafranchian of Europe ( Guérin 1980), all belonging to the genus Dicerorhinus ( McKenna & Bell 1997) View in CoL : the Ruscinian D. megarhinus (de Christol, 1834) , the Late Ruscinian D. miguelcrusafonti Guérin & Santafe, 1978 , the Early Villafranchian D. jeanvireti Guérin, 1972 and the Villafranchian D. etruscus etruscus (Falconer, 1859) . All remains of the Milia rhinoceros correspond to D. jeanvireti .
e genus Dicerorhinus Gloger, 1841 is represented today by D. sumatrensis (Fischer, 1814) : it has many Neogene and Quaternary species in Eurasia and Africa.
Croizet & Jobert (1828: 144-154, pl. I, fig. 7, pl. IV, figs 3-6, pl. V, figs 1-4, pl. VI, figs 1, pl. XI, pl. XII, figs 1, 2, 8) had proposed the name of Rhinoceros elatus for the Auvergne rhinoceros whose remains belonged to a mixture of several species, including D. etruscus and D. jeanvireti , according to the dimensions given in the book of Croizet & Jobert (1828). e exact location of that material is unknown. An important part of it was gathered in Perrier-les Étouaires, where Dicerorhinus etruscus and D. jeanvireti are sympatric. Another part, namely the mandible, which is the first rhino remain described by Croizet & Jobert (1828: 144-146), is from Malbattu, a much more recent site where D. etruscus is the sole rhino ( Guérin, 1980). Such a mixture and the lack of diagnostic characteristics are enough to invalidate the specific name elatus , also completely forgotten for more than a century, but the fact that the Croizet & Jobert’s name is clearly a nomen oblitum is sufficient to discard it.
Dicerorhinus jeanvireti Guérin, 1972 is a very large rhino with slender legs. e massive facial area is very long; the nasals are quite large and reach far forward, bearing an ossified septum in their anterior part. e pseudo-auditory meatus is closed ventrally.
e mandible bears a long symphysis and presents a slightly convex lower edge.
e M1/ and the M2/ have an almost flat ectoloph with a very slight paracone fold; there is always a crochet, rarely a crista; the protocone is constricted, there is no internal cingulum. e P3/ and the P/4 with an almost flat ectoloph and a very small paracone fold, always present a crochet, very frequently a crista and often a closed medifossette; the protocone is not constricted and there is always a lingual cingulum. e long bones are very large and slender, the limb segments 2 (radius, tibia) and 3 (metacarpals, metatarsals) are very elongated, and the median metapodials are long and relatively slender.
e species, defined in Vialette, is also known in other French sites, such as Perrier-Les Étouaires and the Desnes-Vincent-Bletterans region near Dôle.
It is present in the Early Villafranchian of Italy, in Becchi di Castelnuovo Don Bosco (Astigiana), Dusino, Villafranca d’Asti (Piedmont), Capannoli, Montopoli (Lower Valdarno), Pradalbino (Monteveglio, District of Bologna), the Siena region ( Guérin 1972, 1980). More recently Campanino et al. (1994) described in detail beautiful post-cranial material discovered in Roatto, near Villafranca d’Asti.
Dicerorhinus jeanvireti is probably present in the Netherlands, in the Scheldt Estuary ( Hooijer 1981) and in Linden ( Hooijer 1985). It is quoted from Germany in the Hambach lignite mine, West of Cologne ( Lacombat & Mörs, 2008), a site no farther north than the two preceding ones. It has been recognized in Czechoslovakia, in Hajnacka ( Fejfar 1964; Durisova 2004) and in Strekov ( Schmidt & Halouzka 1970). It was also reported in Bulgaria, in Musselievo (Late MNQ 15 following Spassov 2005), a site which may be the oldest locality of the species.
Finally, in Romania Radulescu et al. (2003) reported “ Stephanorhinus elatus ” or “ S. cf. elatus ” in the Brasov Iaras – 2 Depression (MNQ 16a), in Araci-Fagului Fantana (MNQ16a), in Ilieni Basin (MNQ 16) and Cernatu (MNQ 16), and in the Dacian Basin, in Covrigi (MNQ 16a), Groserea (MNQ 16a) and Cernatesti (MNQ 16a), all three in Oltenia.
Lacombat & Mörs (2008) pointed out that the Early Villafranchian rhinos of Kvabebi ( Georgia), and of Udunga and Nizhnavodyanoy ( Russia) could be D. jeanvireti .
Dicerorhinus jeanvireti is characteristic of the Early Villafranchian (zone MNQ 16).
MATERIAL
Most of the 1685 fossils from Milia belong to Mammut borsoni . 197 pieces are attributed to rhinoceros; 144 are bones and bone fragments, 35 are isolated teeth and tooth fragments, 16 are mandibles and mandible fragments with teeth, a maxilla fragment with upper cheek teeth and two skulls: an almost complete one with complete dentition and a cranial dome. From the postcranial skeleton very few bones are complete, while 68 specimens are measurable and listed as follows:
– two skulls: a skull with complete dentition MIL 162, which is associated with the right hemimandible fragment and a facially incomplete skull MIL 1501 without its teeth;
– a fragment of right maxilla MIL 1476, with P2/ damaged and P3/;
– twelve complete or fragmentary mandibles: a mandible with right P/2-P/4 and left P/3-M/1 MIL 658; a mandible with complete tooth rows SGP 1; a right hemi-mandible with M/2-M/3 MIL 244; a left hemi-mandible fragment on the skull associated with M/2-M/3 MIL 162a; a right hemi-mandible with full dentition MIL 281; a right hemi-mandible fragment with M/2-M/3 MIL 245; a right hemi-mandible fragment with M/3 incomplete MIL 1477; a fragment of ramus MIL 267; a right hemi-mandible fragment with D/1 and D/2 MIL 425; a left hemi-mandible fragment with P/2 and P/3 incomplete and P/4 MIL 245; a right hemi-mandible fragment with P/4 and fragments of molars MIL 1324; a right hemi-mandible fragment with M/1 MIL 1478;
– an isolated P3/ MIL 806 sin; three isolated P4/: MIL 868 dex, MIL 880 sin and MIL 1287 dex; two isolated M1/ MIL 614 sin and MIL 1569 dex; an isolated M2/ MIL 1197 sin; an isolated M3/ MIL 1119 dex; an isolated M/1 MIL 837 dex; two isolated M/3 MIL 1145 dex and MIL 1343 sin;
– a right scapula fragment MIL 205;
– three distal humerus fragments: MIL 673 dex, MIL 751 dex and MIL 841 sin;
– seven radius fragments: MIL 611 dex (proximal half); MIL 656 sin (distal epiphysis); MIL 657 sin (proximal two-thirds); MIL 987 sin (proximal epiphysis); MIL 1315 sin (two-thirds proximal); MIL 1387 dex (proximal third); MIL 1541 sin (proximal third);
– two ulnae: a complete GSP 2 (sin) and an incomplete MIL 1582 (proximal fragment);
– a magnum MIL 758 dex (incomplete);
– two Mc III: a complete MIL 1587 dex and an incomplete MIL 902 sin (proximal half);
– four femur fragments: MIL 613 (caput), MIL 813 (caput), MIL 651 dex (distal fragment) and MIL 811 (distal fragment);
– two patellae: MIL 340 dex and MIL 1645 sin. – five complete or fragmentary tibias: MIL 819 dex (complete); MIL 612 dex (distal two-thirds); MIL 1582 sin (distal fragment); MIL 1642 sin (distal fragment); MIL 1645 sin (distal fragment). – four astragali: MIL181 dex, MIL 367 sin, MIL 852 sin, MIL 1192 (fragment);
– a complete calcaneum MIL 1703 sin and a calcaneum fragment MIL 723 sin;
– a complete cuboid MIL 1665 dex;
– two Mt III: MIL 368 dex (complete) and MIL 710 sin (proximal fragment);
– two Mt IV: MIL 204 sin (complete) and MIL 767 dex (proximal fragment);
– three distal metapodial fragments, undifferentiated: MIL 741 (axial), MIL 1407 (axial), MIL 1440 (abaxial);
– three phalanges: MIL 206 (Ph I axial, complete), MIL 347 (Ph I abaxial, complete), MIL 1385 (Ph III, fragment).
Sesamoids have not been studied, nor have vertebrae and ribs, or small fragments.
e entire sample is attributed to Dicerorhinus jeanvireti .
e most abundant anatomical specimen represented among the Milia rhino material is the mandible, which consists of two sub-complete adult specimens and nine more or less fragmentary hemi-mandibles; seven right (including a juvenile) and two left ones. Radius follows, seven of which are studied: five left (four of which are proximal fragments) and two right (both proximal) ones, whose matching remains are doubtful.
In conclusion, among the Milia material there are at least eight rhino individuals, adults or subadults and a juvenile one.
DESCRIPTION e reference material used for the Simpson diagrams is the average of 30 to 40 adult individuals of the recent Diceros bicornis (Linnaeus, 1758) , the African “Black” Rhinoceros (values in Guérin 1980, plus a few new specimens).
Skull e “wedge-shaped” general form of the Rhinocerotidae skulls is evident in inferior and superior view ( Fig. 3A, B View FIG ). In profile view ( Fig. 3 View FIG C-E), the relative length of the massive facial area is great, which is characteristic of the genus and the species: the ratio of the distances outer occipital bump-ante-orbital process (variable 8 in Table 2 [see Appendices]) / occipito-nasal length (variable 1) reaches 0.57 and 0.56 for the two Milia skulls when the average values are 0.556 for D. megarhinus and 0.55 for D. etruscus etruscus . e elevation of the massive major occipital crest is well marked; it overhangs the occipital condyles behind. e tip of the nasals extends forwards to the tip of the premaxillae as in the Vialette specimens; the ossified nasal septum extends forwards to the tip of the nasals. On the skull MIL 162 the ossified nasal septum is more than 121 mm in length (it is broken further back), its maximum thickness (back and base of the posterior fracture) reaches 11.5 mm. e skull MIL 1501 shows no trace of ossified nasal septum, but the absence of teeth does not allow the estimation of the age for this individual, which might not be fully adult; note also that enius (1955) suggested that there are a few adult specimens of D. etruscus in which this wall is not ossified probably representing females.
e nasal septum is never ossified in D. megarhinus .
e post-glenoid apophysis is very wide from front to rear, more vertically developed than the post-tympanic and paroccipital apophyses and it is tilted forward and down ( Fig. 3D View FIG ). e posttympanic apophysis is very short. e paroccipital apophysis is quite long, sub-vertical, and bends slightly forwards at the end. In the skull MIL 162, the level of the posterior end of the nasal notch is between P3/-P4/; this position varies from the middle of P3/ to the interval P3/-P4/ in the two specimens of D. jeanvireti from Vialette. e infraorbital foramen is above the middle of P4/, it is more anterior in Vialette (interval P3/-P4/ in front of P4/). Finally, the level of the anterior border of the orbit is above between M2/ and M3/, it is above the midst of M2/ on the only Vialette skull where it is observable. Antoine (2002) showed that generally in the family Rhinocerotidae , evolution is reflected by a receding of these relative positions. In superior view ( Fig. 3B View FIG ), the insertion of the posterior horn (frontal) is very broad, and that of the anterior one (nasal) is well marked. e front of the nasal bone is rounded with a small recess in the sagittal plane. e major occipital crest of the skull MIL 1501 is greatly depressed in the middle, while the skull MIL 162 is less concave in its central part. e ante-orbital processes are strong, the supra-orbital ones are slightly prominent and the post-orbital ones are very low.
e occipital ( Fig. 3 F View FIG ) is nearly vertical in its lower and middle parts, and tilted slightly backwards and upwards in its superior part. e depression in the centre of the major occipital crest is clearly visible from this angle. In ventral view ( Figs 3A View FIG ; 4A View FIG ), the basioccipital muscular tubercule is strong. For the skull MIL 162 the posterior part of the post-palatal notch is posterior to M2/ and it is more advanced (interval M1/-M2/) in the only Vialette skull where such an observation is possible.
e skulls of the Milia rhinos clearly differ from those of D. megarhinus : the latter especially possess thicker and wider nasals which are free at their rostral end, whithout any ossified septum, and a longer nasal notch; the major occipital crest is less depressed medially and with a small central bump; shape and relative lengths of the postglenoidal, posttympanical and paroccipital apophysis are different.
e dimensions are given in Table 2 (see Appendices). All measurements exceed those of D. etruscus and are slightly lower than those of D. megarhinus . Most of them are close to those of the two skulls of Vialette D. jeanvireti . e latter, although of similar general proportions, are, however, smaller than those from Milia: in Vialette they are shorter, their nasal notch is shorter, their cranial dome is narrower; the rear of the nasal notch, the infraorbital foramen, the anterior border of the orbit and the rear of the post-palatinal notch are all more reduced in Milia than in Vialette. Due to the small sample available, the only Plio-Villafranchian rhino of Europe in which evolutionary trends have been highlighted in the skull is Dicerorhinus etruscus etruscus : they particularly include the increasing distance of the nasal notch-orbit and the enlargement of the cranial dome ( Guérin 1980). It is very likely that these trends are similar to D. jeanvireti , which belongs to the same subgenus, in which case the Milia specimens are more advanced and therefore somewhat more recent than those of Vialette.
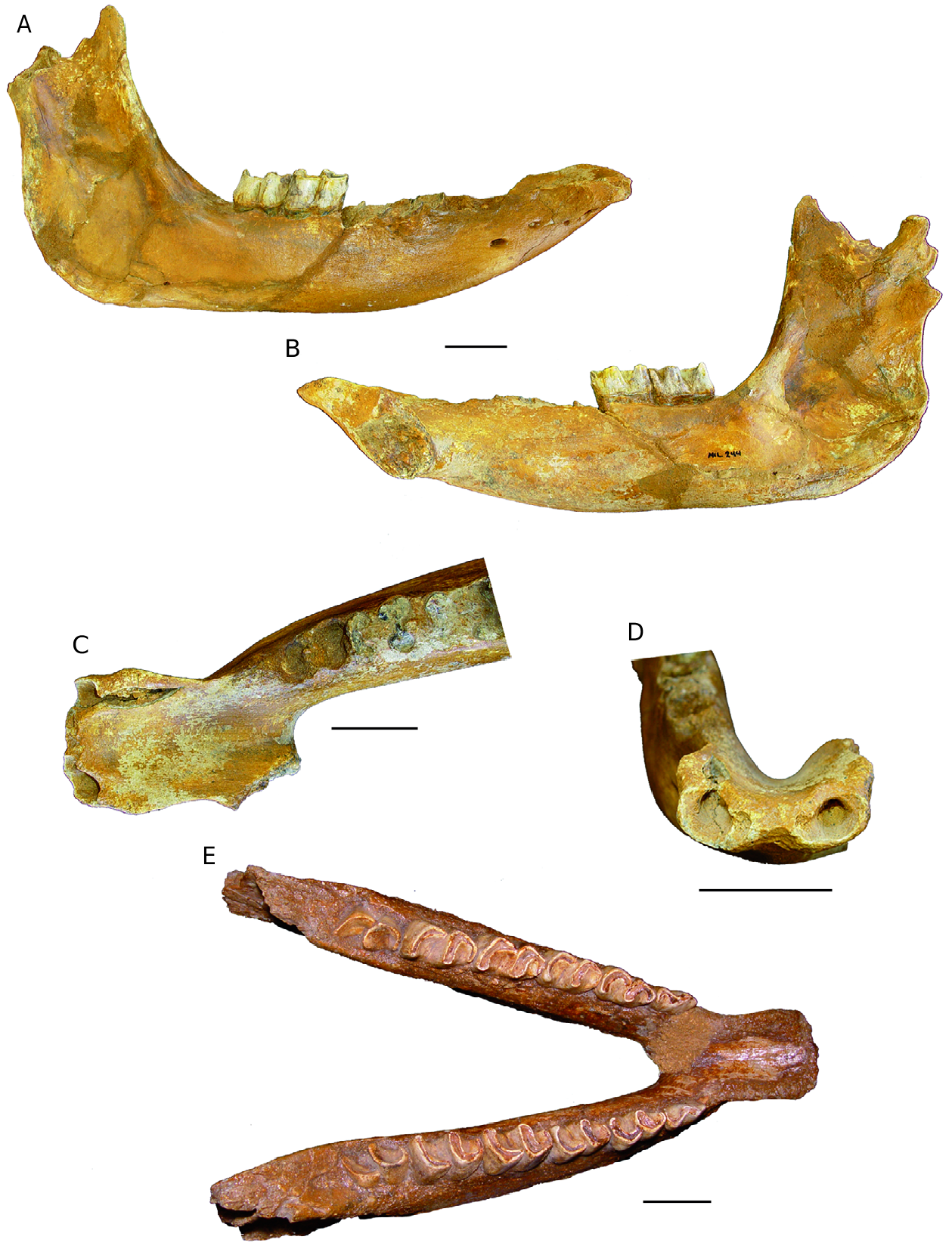
Mandible e ventral edge of the horizontal branch (corpus) is straight in profile view ( Fig. 5A, B View FIG ). e mandibular foramen is at the alveolar border. e scalloped talon is very prominent backwards, a little beyond the posterior edge of the condyles.
e anterior border of the ramus is subvertical, barely oblique backwards and upwards, in its upper part.
In superior view the symphyseal region ( Fig. 5C, E View FIG ) shows the slightly spatulate space, observed in D. megarhinus and D. jeanviret i ( Guérin 1980). e ventral side of the symphysis bears no median crest.
e specimen MIL 244 bears on the front border of the symphysis two small alveoli for I/1, and two large alveoli for I/2, which were well developed into small tusks ( Fig. 5D View FIG ). e spacing between the two tusks is 27 mm and the right alveolus (the only well preserved one) has DT = 19 mm for DDV = 15 mm.
e alveoli of the tusks are smaller for MIL 658 (DT = 10 mm, DDV = 9 mm) and less far apart (20.5 mm); these external alveoli, especially for MIL 244, are larger than those typically observed in D. jeanvireti . Altough not functional as were the very large I/2 of the Miocene Dicerorhinus , which were powerfull weapons, the down-sized I/2 of D. megarhinus , button-shaped with a diameter of about 1 cm and a length of about 2 cm, are clearly larger than those of D. jeanvireti .
On the mandible GSP 1 the posterior edge of the symphysis is in the middle of P/2, on MIL 658 it is in the middle of P/3 and on MIL 281 it is in the rear of P/3; the position being more distant than in seven specimens of D. jeanvireti , where it further forwards ( Guérin 1980): it ranged from the mid-P/2 to the extreme front of P/3. Antoine (2002) showed that among Rhinocerotidae , the relative position of the posterior border of the symphysis usually tends to recede during the evolution of the family.
e dimensions of the mandibles are given in Table 3 (see Appendices). ey are all in the range of variation of 2 to 14 specimens of D. jeanvireti , while those of 2 to 42 specimens of D. etruscus etruscus are smaller and those of 1 to 42 specimens of D. megarhinu s are a little larger.
Although of more or less similar dimensions the mandibles of the Milia rhinos clearly differ from those of D.megarhinus : the latter especially possess a more elongated symphysis, a longer, thicker and lower corpus, a talon more proeminent backwards and an anterior border of the ramus oblique backwards and upwards.
Upper teeth
An anterior upper tooth is preserved on the right side of the skull MIL 162 ( Fig. 3E View FIG ). is is the first time that an upper front tooth is seen in Pliocene and Pleistocene rhinoceroses of Europe. Conical, high, it goes just beyond the base of the premaxilla and is located at the base of this bone, 76.5 mm in front of P2/ in the same side, and 54 mm from the rostral end of the intermaxillary; it is probably a dI/2 or I/2, the inability to assess the thickness of the enamel prevents its determination as a deciduous or permanent tooth. Its transversal diameter is 9 mm, the antero-posterior diameter is 10 mm and its height reaches 22.5 mm. is incisor does not show any obvious trace of wear and lacks occlusion, thus it was not functional.
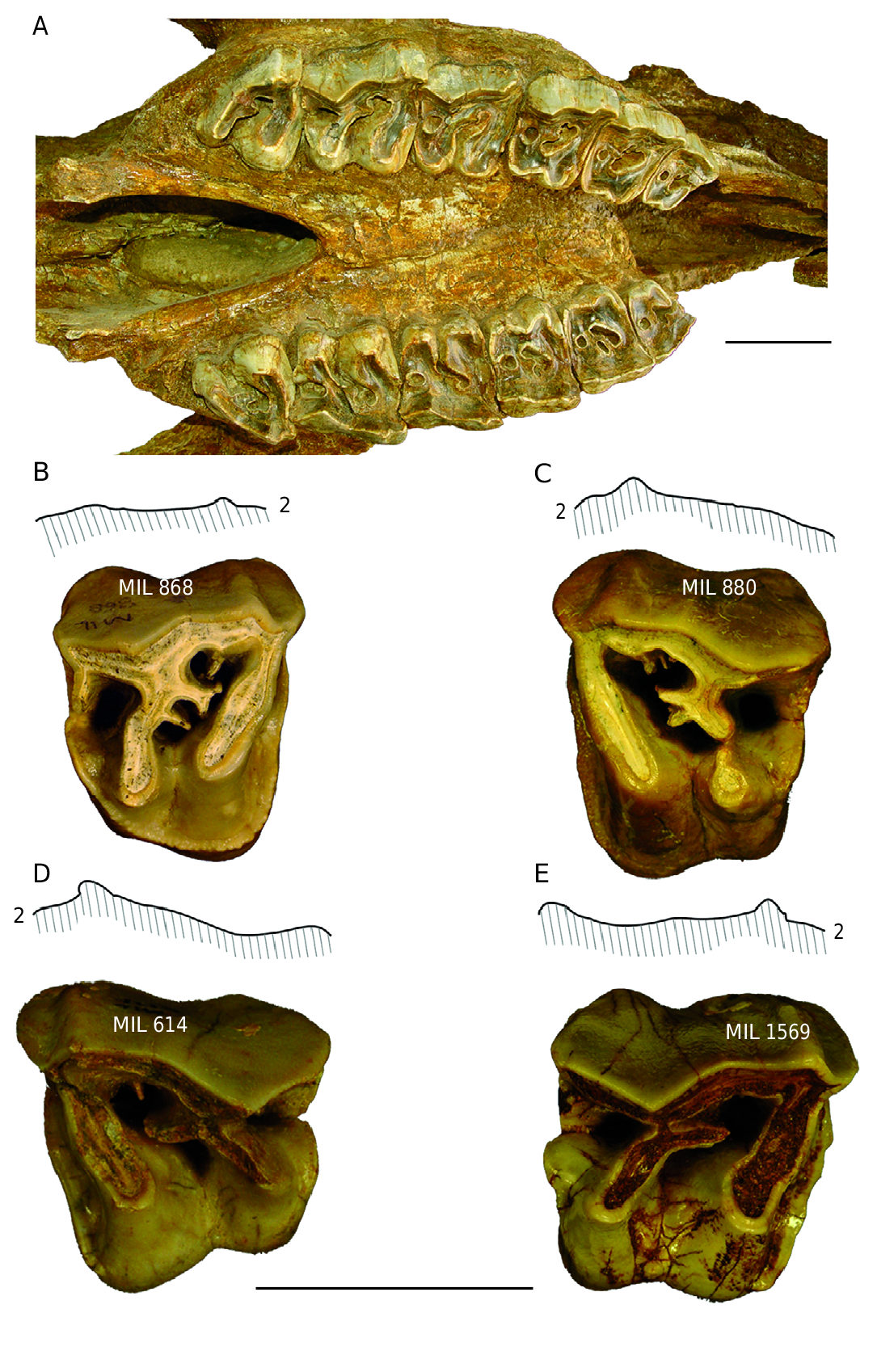
e cheek teeth of the skull MIL 162 ( Fig. 4A View FIG ) are all present but worn.
e protoloph of P2/ is notched in the middle, while the only internal fold is a crista and there is a continuous lingual cingulum.
On the P3/ ectoloph there is a strong parastyle, also there is a small paracone fold and a very small metacone fold. Crista and crochet are present and on the left tooth, the two folds merge distally to form a closed middle fossette. e lingual cingulum is discontinuous. On maxilla fragment MIL 1476, the P3/ has no crista. e unworn isolated P3/ MIL 806 has a multiple crochet and a small crista, no constriction of protocone or lingual cingulum and there is a small mesostyle. e height of the crown is 43.5 mm and has a hypsodonty index of 101.2.
e P4/ is constructed along the same lines as the P3/, but the crochet is multiple, the lingual cingulum is discontinuous and the protocone is slightly constricted. A very small mesostyle is noted in the middle of the ectoloph. e isolated P4/ MIL 868 ( Fig. 4B View FIG ) is characterized by its multiple crochet, its lack of constriction of protocone and its continuous lingual cingulum; the isolated P4/ MIL 880, also bears a multiple crochet but not lingual cingulum, and the hypocone is isolated at its end ( Fig. 4C View FIG ); finally the P4/ MIL 1287 shows a continuous lingual cingulum.
e M1/ has an ectoloph, whose posterior part is directed outwards and a shows a strong paracone fold A crochet is the only internal fold and there is no lingual cingulum; the constriction of the protocone is intense. e M1/ MIL 614 ( Fig. 4D View FIG ) is characterized by a discontinuous lingual cingulum, a slight crista in addition to its crochet, and a small mesostyle. Another isolated M1/ MIL 1569 shows a moderately developed constriction affecting the hypocone as well as the protocone, and a lingual cingulum reduced to two buttons ( Fig. 4E View FIG ).
e M2/ has the same ectoloph profile as in M1/. ere is a crista and a crochet. Constriction of the protocone is strong. ere is no lingual cingulum. e isolated M2/ MIL 1187 is very similar but lacks the crista.
e M3/ has no crochet and a discontinuous lingual cingulum. Constriction of protocone is well marked. e isolated M3/ MIL 1119 is very similar.
e morphology of the Milia upper cheek teeth corresponds to that of D. jeanvireti , although in the molars the paracone fold is more marked and the ectolph less flat than for the molars of Vialette.
e dimensions of the upper cheek are given in Table 4 (see Appendices). In the Milia specimens the dimensions are slightly higher than those of the comparative sample of D. jeanvireti .
e four Plio-Villafranchian European rhino species pertaining to the same genus, there are no important differences in the morphology of their upper cheek-teeth; the ectoloph profiles are only slightly different, e.g., a thicker paracone fold for the D. megarhinus P3/-4/.
Lower teeth e type of construction of the lower cheek teeth of Pliocene and Pleistocene rhinoceroses is very monotonous in general, and there are no anatomical characteristics that distinguish the diverse species of Dicerorhinus . e lower cheek teeth of the Milia rhinoceros ( Fig. 5E View FIG ) can only be specifically identified from their dimensions.
e M/2 of the mandible MIL 162 bears two Vshaped lingual valleys, which between them present a small difference in level something that happens in most cases of the M/2 of D. jeanvireti , however some of them have an anterior U-shaped valley.
e M/3 of the mandible MIL 162 with its anterior V-shaped valley, its posterior U-shaped valley and the high difference of the level between the two valleys, also represents the general case, the variability focuses essentially on the difference in level between the two valleys. It is confirmed in the Milia specimens, that the two isolated M/3s MIL 1145 and MIL 1343 have their two valleys V-shaped, with a difference in level which is reduced in the former but stronger in the latter.
In D. jeanvireti the P/2 does not show any particular character, the P/3 and P/4 lack of lingual or buccal cingula and have V-shaped valleys with a difference in levels between them from low to medium. ese observations apply to the lower premolars from Milia.
e dimensions are given in Table 5 (see Appendices). e Milia lower cheek teeth fall within the range of variation of D. jeanvireti , some being marginally larger than the maximum values of the comparative sample.
Radius and Ulna ere are six specimens of proximal radius, of which specimen MIL 657 is the best preserved ( Fig.6A, B View FIG ). e anatomical characteristics are those of D. jeanvireti , particularly the proximal articulation distinctly undulating on its anterior border, with a strong medial re-entrant, to the posterior edge forming an obtuse angle, and on the lateral facet slightly extending outwards, whose front edge is recessed from the front edge of the medial facet ( Fig. 6A View FIG ). In anterior view, notable are the strong lateral tuberosity of the proximal epiphysis, which is well developed below the articulation, and the convex medial border of the same epiphysis, and the strong insertion of the biceps brachii shifted slightly medially ( Fig. 6B View FIG ).
e proximal DT varies from 97.5 to 102 mm (average for four specimens: 99.6 mm), the DAP proximal 64 to 75mm (average for four specimens: 70.8 mm), the DT of diaphysis 57 to 60 mm (average for two specimens: 58.5 mm), the DAP diaphysis 37.5 to 39 mm (average for two specimens: 38.3 mm).
ese dimensions fall within the range of variation of a sample of 10 to 12 radii of D. jeanvireti , whose respective average values are 101.6 mm, 69.9 mm, 56.5 mm and 39.1 mm ( Guérin 1980: 479, table92).
e single almost complete ulna ( Fig. 6C View FIG ) measures 560 mm long, which is much more than the maximum value measured on a sample of five ulnae of D. jeanvireti (maximum 529 mm, with an average of 519.4 mm), but less than 580 mm in the Roatto ulna ( Campanino et al. 1994).
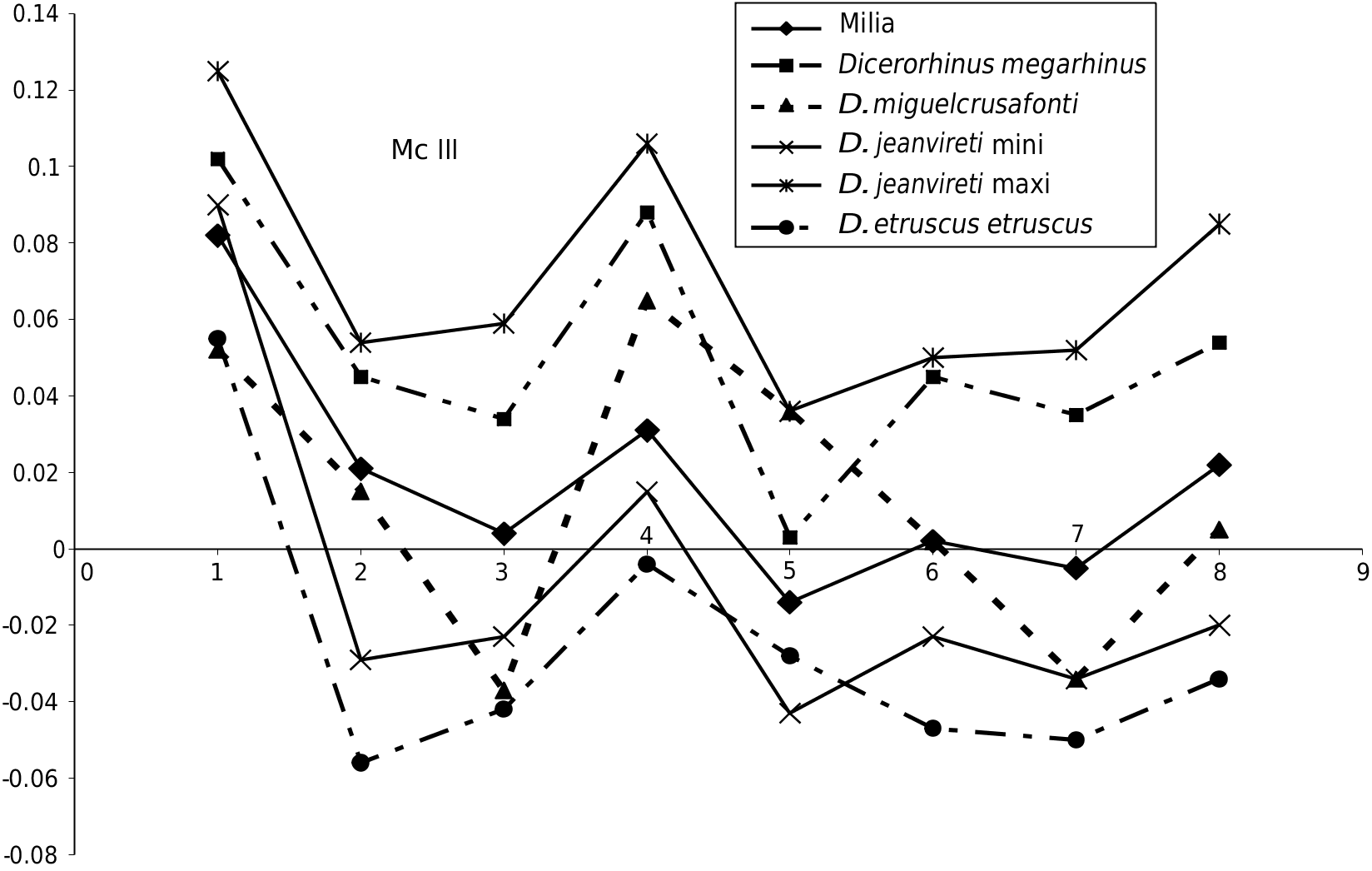
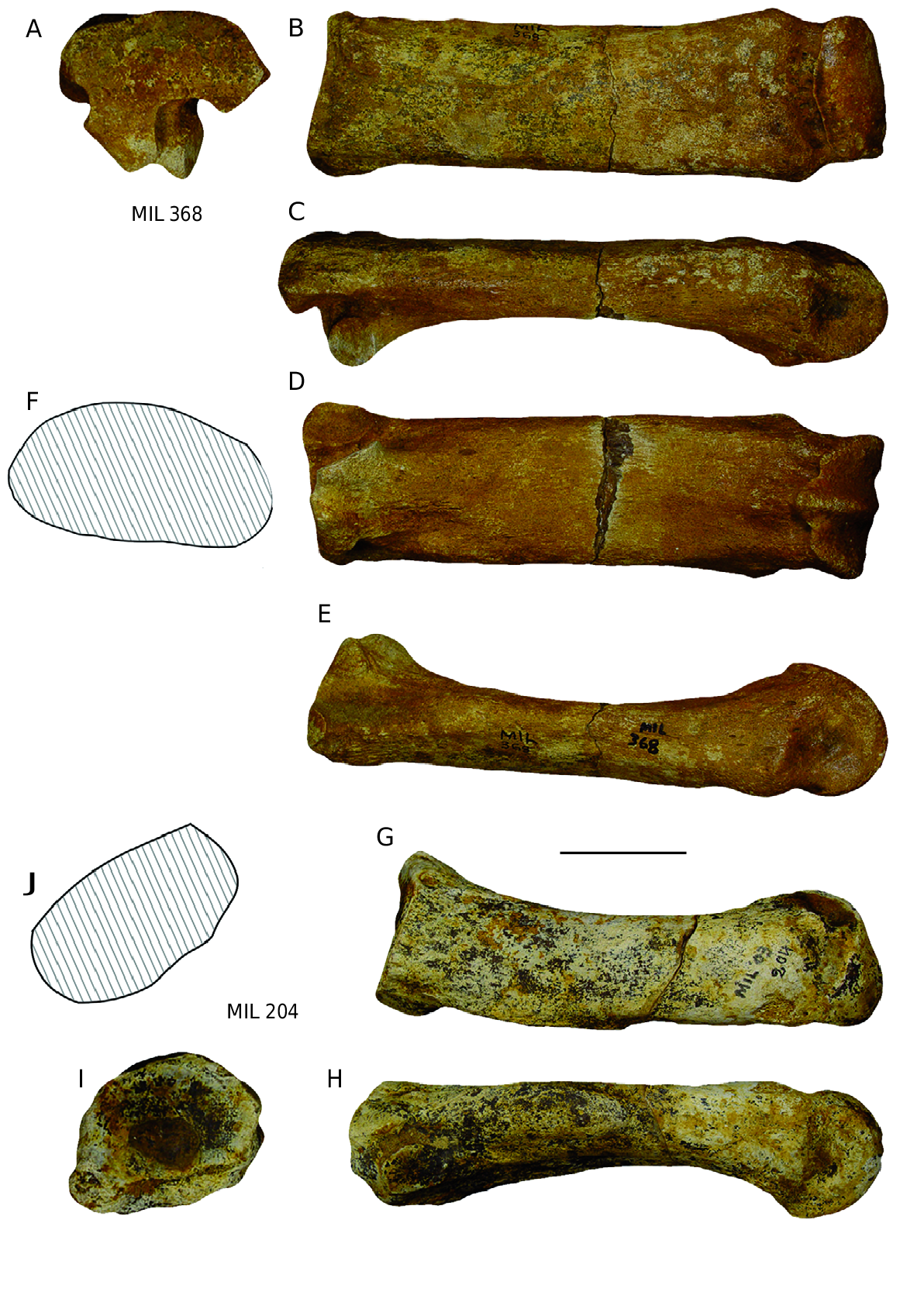
Mc III ere is a complete third metacarpal ( Fig. 7 View FIG ) and a half one. e anatomical characteristics are those of D. jeanvireti : a long and relatively slender bone ( Fig. 7A, F View FIG ); a proximal articulation with a clearly convex anterior border ( Fig. 7C View FIG ); an anterior proximo-lateral articular surface much larger than the posterior, with a distal part as high as the proximal part; a proximo-lateral posterior facet a little higher than wider ( Fig. 7B View FIG ), which is noted to be of trapezoidal shape instead of triangular as in Vialette; a diaphysis of elliptical midsection and slightly depressed in the posterior border ( Fig. 7D View FIG ).
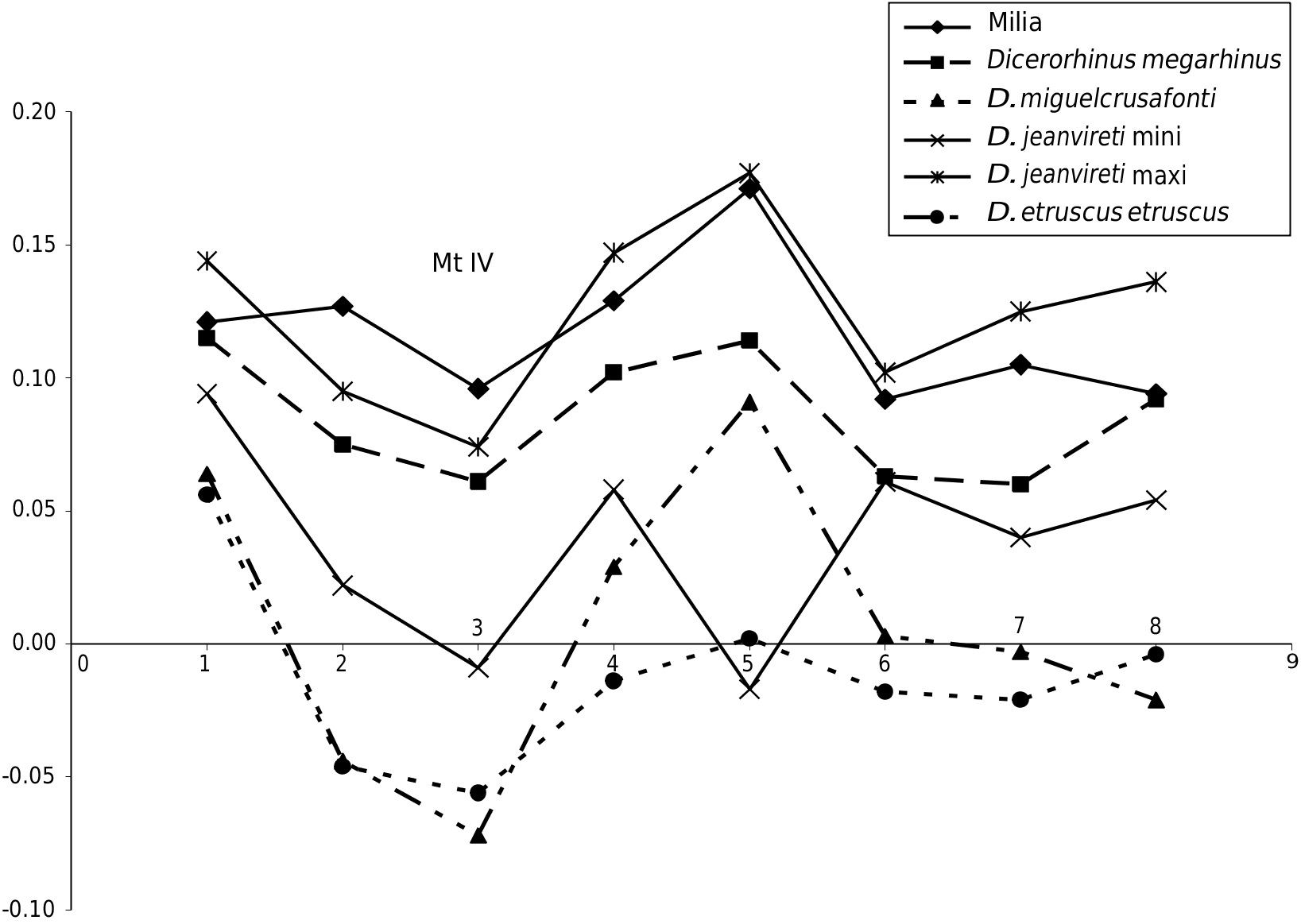
e dimensions are given in Table 6 (see Appendices). Except for the length, which is somewhat smaller than the minimum value of the sample of comparison,the Milia specimens fall within the range of variation of 8 to 11 specimens of D. jeanvireti Mc III. e Simpson diagram in Figure 8 View FIG shows that the extreme values of D. jeanvireti surround those of Milia rhinoceros, except for the length.
e peculiar design of the graph corresponding to D. miguelcrusafonti was noted incidentally.
In 1980, Guérin was not able to identify the evolutionary trends on the D. jeanvireti Mc III.
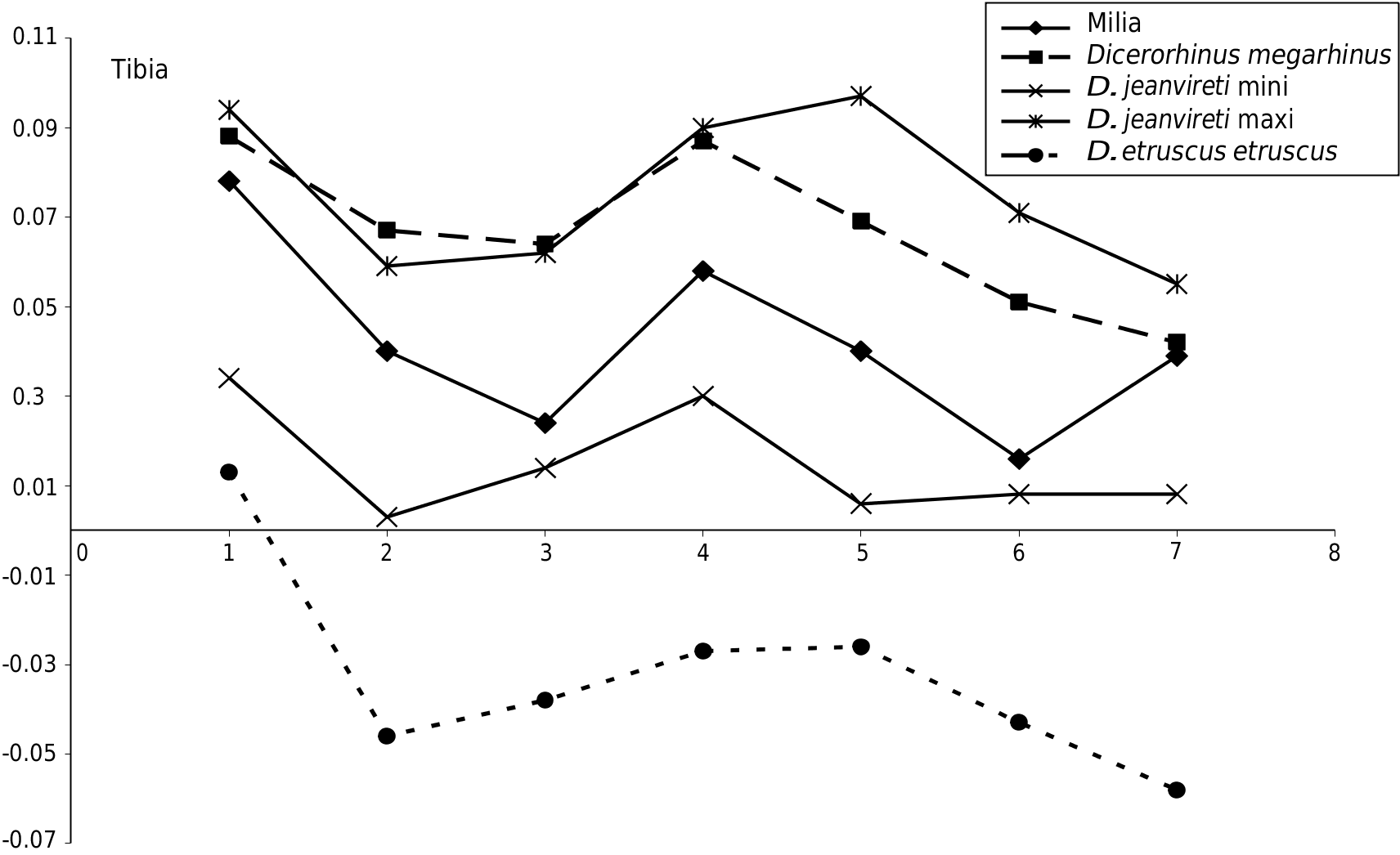
Tibia e single complete Milia specimen MIL 819 ( Fig.6 View FIG D-F) shows the anatomical features of D. jeanvireti , which were established by Guérin in 1972 and 1980: proximal epiphysis with strong tibial tuberosity untrammelled and projecting upwards; brims of the tibial spine shifted backwards (the lateral being more posterior), but not really shifted vertically, the popliteal notch strongly carved; distal epiphysis with the medial malleolus very clear and rather square in outline and relatively low fibular notch.
e distal articulation has DT = 85 mm for DAP = 64 mm; the DT distal articulation of a Desnes tibia reaches 86 mm. e two diameters of articulation are stronger in the Roatto specimen: 92.5 and 75 mm ( Campanino et al. 1994).
e other dimensions are given in Table 7 (see Appendices). e dimensions of Milia tibiae are within the range of variation of 8 to 13 specimens of D. jeanvireti , and are close to the average of this sample. e Roatto tibia is significantly longer ( Campanino et al. 1994).
e Simpson diagram in Figure 9 View FIG shows that the complete Milia tibia is perfectly framed by the extreme values of D. jeanvireti . It is also noted that the proportions of D. megarhinus are rather different compared to the maximum values of D. jeanvireti ; the average length is a little lower, but the proximal epiphysis is larger, the diaphysis is less deep and the distal epiphysis is smaller.
In 1980, Guérin showed that from Vialette (the dawn of zone MNQ 16) to Montopoli (near the end of the zone), the tibia of D. jeanvireti seems to have tendencies of elongating and reducing the width of the proximal epiphysis.
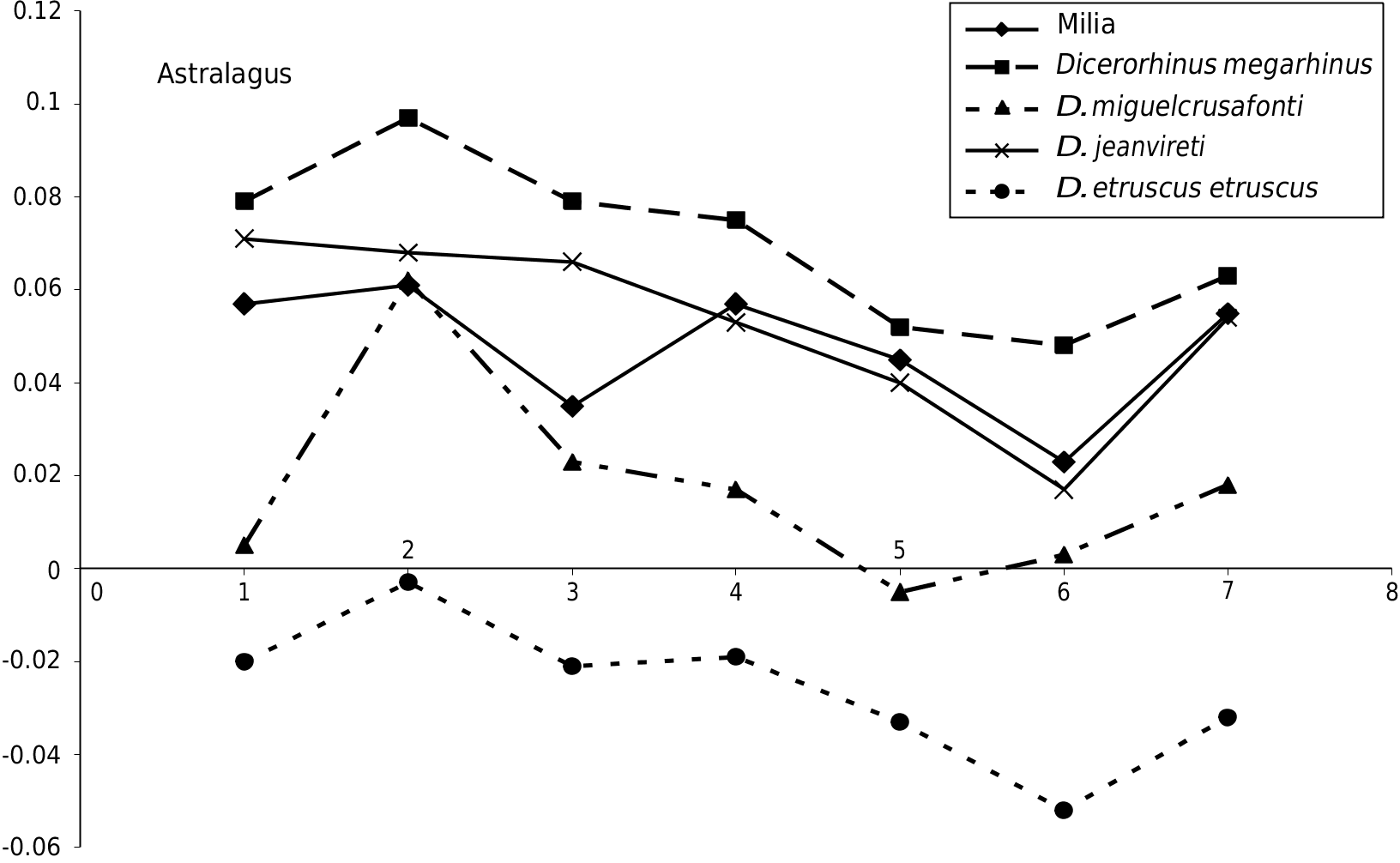
Astragalus e trochlea of the astragalus in D. jeanvireti is wider, shallower and more asymmetrical than in D. megarhinus ; this is also observed in Milia but the trochlea is carved and more symmetrical on MIL 367 and MIL 181 specimens.
In medial view, the distal tubercle is shifted backwards and near the distal edge of the bone.
In distal view, the articular surface for the navicular has its anterior border depressed; in relation to this anterior border, the articular surface for the cuboid is slightly shifted forwards.
All these characteristics are those of D. jeanvireti .
e dimensions are given in Table 8 (see Appendices). e dimensions of the three Milia astragali are within the range of variation of 8 to 15 astragali of D. jeanvireti . e Roatto astragalus is significantly larger ( Campanino et al. 1994).
Figure 10 View FIG confirms the proximity of the average size of Milia with the average of D. jeanvireti ; compared to D. megarhinus the width is similar but the DAP and particularly the height are lower. Note incidentally an analogy between the graphs corresponding to D. miguelcrusafonti and D. etruscus etruscus , which reveals a certain similarity between the proportions despite the much greater size of the former.
e unique evolutionary trend found for the astragalus between the beginning and the end of the zone MNQ 16, is the significant reduction of the transversal diameter ( Guérin 1980).
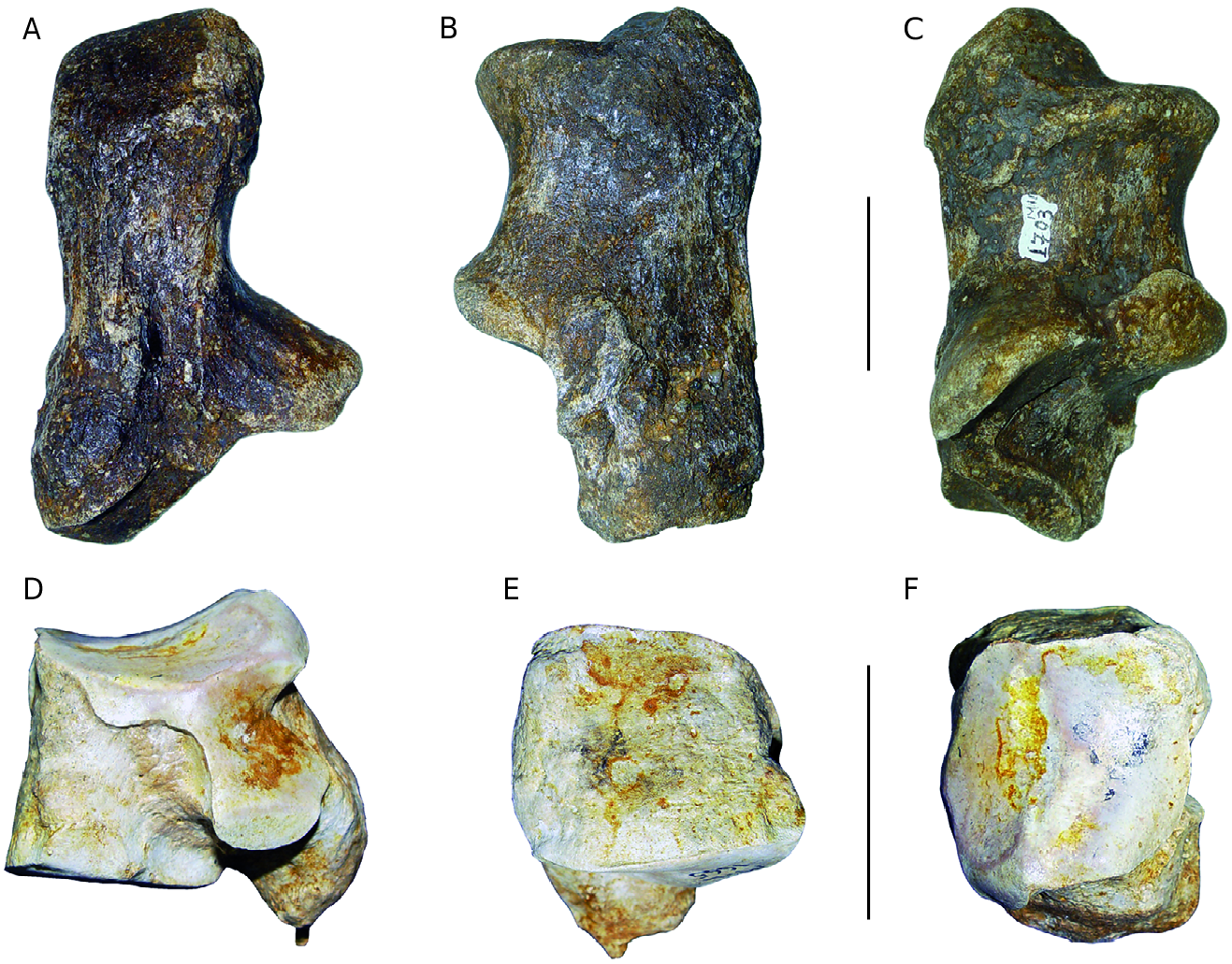
Calcaneum
In posterior view ( Fig. 11A View FIG ), the sustenculum tali axis is slightly inclined towards the bottom and it makes a slightly obtuse angle with the axis of the body of the bone. In lateral and medial views ( Fig. 11B, C View FIG ), the front of the tuberosity is situated slightly behind the beak (= foremost part of the bone), the anterior edge between these two points is concave. e posterior edge of the lateral face is rectilinear for the lower three quarters of its height. e dimensions of the Milia calcaneum are situated well within the range of variation of 10 and 14 of D. jeanvireti calcanea (Appendices, Table 9).
Cuboid e medial face ( Fig.11D View FIG ) is typical of D. jeanvireti .
e anterior face ( Fig. 11E View FIG ) is trapezoidal and not taller than broader, with a medial edge longer than that of the medial face. On the proximal articulation ( Fig. 11F View FIG ) the lateral facet is shorter but once again broader than the medial one.
e dimensions (Appendices, Table 10) of the Milia cuboid are situated within the range of variation of two to nine cuboids of D. jeanvireti , somewhat below the mean except for the height of the bone.
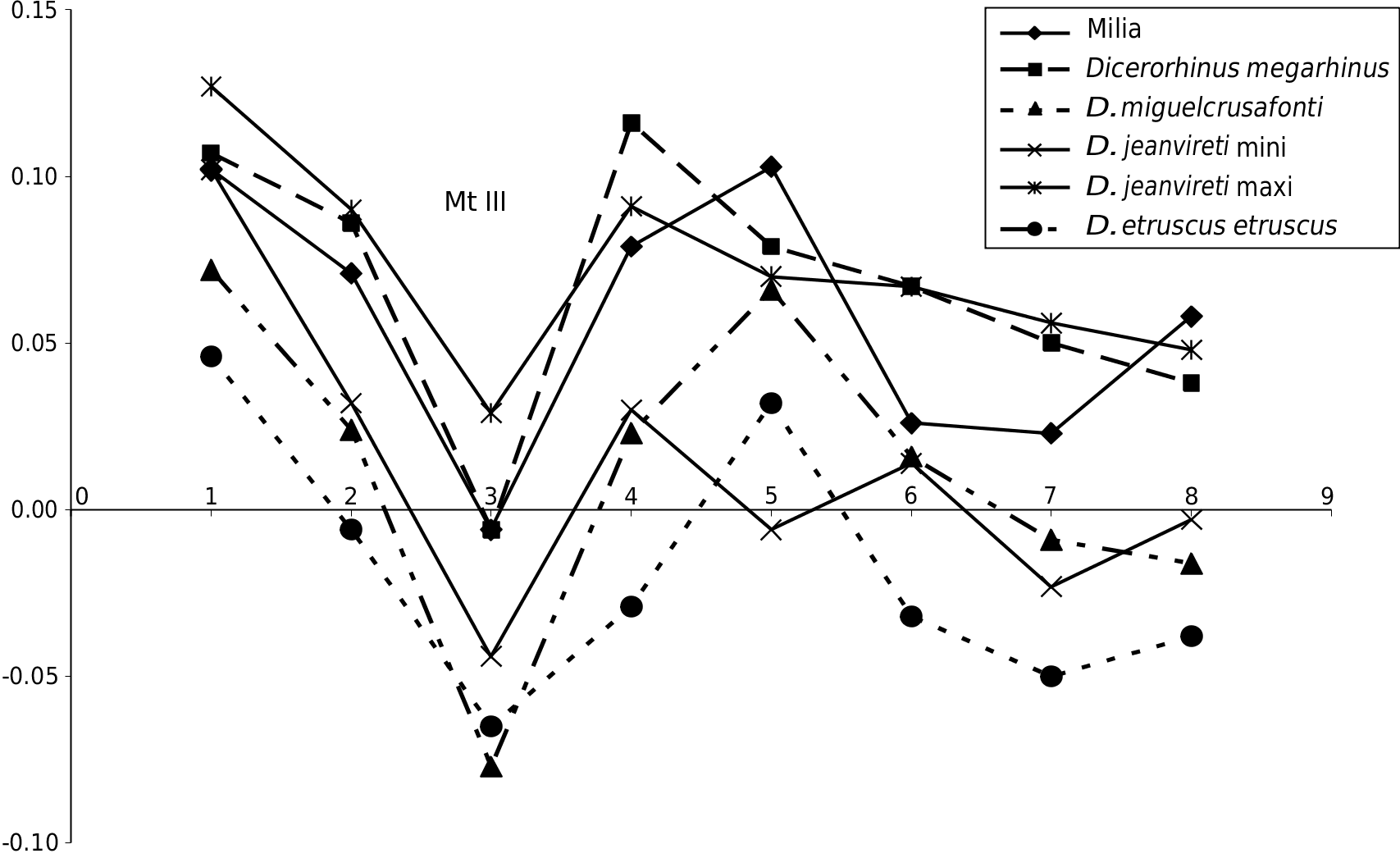
Mt III e complete Mt III ( Fig. 12 View FIG A-F) presents all the anatomical characteristics of D. jeanvireti : an elongated and relatively slender bone ( Fig. 12B, D View FIG ), the proximal articulation is not very broad but deep and the anterior border is regular and weakly convex ( Fig.12A View FIG ); the proximo-lateral articulation consists of two facets with similar length and height, the former being well above the posterior ( Fig. 12C View FIG ).
e proximo-medial articulation has two distinct facets in which the posterior one, situated below, is significantly larger than the anterior one ( Fig. 12E View FIG ).
e cross section of the diaphysis is trapezoidal with a rounded posterior border, which is slightly depressed ( Fig. 12F View FIG ).
e dimensions are given in Table 11 (see Appendices). e Milia specimens fall in the range of variation of 6 to 11 specimens of Mt III of D. jeanvireti , two of them (the DT of diaphysis and the distal articulation) are, however, marginally higher than the maximum values of the sample for comparison. e Roatto Mt III is significantly larger ( Campanino et al. 1994).
e Simpson diagram in Figure 13 View FIG shows that the extreme values of D. jeanvireti frame more often those of the Milia rhinoceros, where however, the DAP of diaphysis and the distal epiphysis exceed the maximum values observed. Note here again a certain parallelism in the graphs corresponding to D. miguelcrusafonti and D. etruscus etruscus .
From the beginning to the end of the zone MNQ 16, the proximal epiphysis tends to shrink; the diaphysis becomes shallower and narrower at its base ( Guérin 1980).
MtIV e complete specimen represented in Figure 12G, J View FIG , also presents the anatomical and biometric characteristics of Dicerorhinus jeanvireti . e bone is slender and elongated ( Fig. 12G, H View FIG ). e proximal articulation, a little wider than longer, is pentagonal, with an anterior border having a very open angle and a posterior margin barely emarginated ( Fig. 12I View FIG ). e proximo-medial articulation bears two facets, the anterior one, which is smaller and more or less trapezoidal in shape and is located much higher than the posterior one, which is larger and elliptical ( Fig. 12H View FIG ).
e cross section of the diaphysis ( Fig. 12J View FIG ) is a rounded parallelogram, with the medial border shorter than the lateral one.
e dimensions are given in Table 12 (see Appendices). e Milia specimen is within the range of variation of a sample of five to ten specimens of D. jeanvireti , but its proximal epiphysis has two diameters slightly greater than the maximum values of our sample. Durisova (2004) gives the dimensions of two Mt IV which were collected during new excavations in Hajnacka. ese dimensions, reported in Table 12 (see Appendices), correspond perfectly to those of D. jeanvireti .
Figure 14 View FIG shows that the proportions of the Milia Mt IV are those of D. jeanvireti , with dimensions close to the maximum values of the comparative sample, and confirms the high volume of the Milia proximal epiphysis. e evolutionary trends ( Guérin 1980) are: an increase in length, a narrowing of the diaphysis at mid-shaft and a broadening to its base.
CONCLUSION OF THE ANATOMICAL STUDY
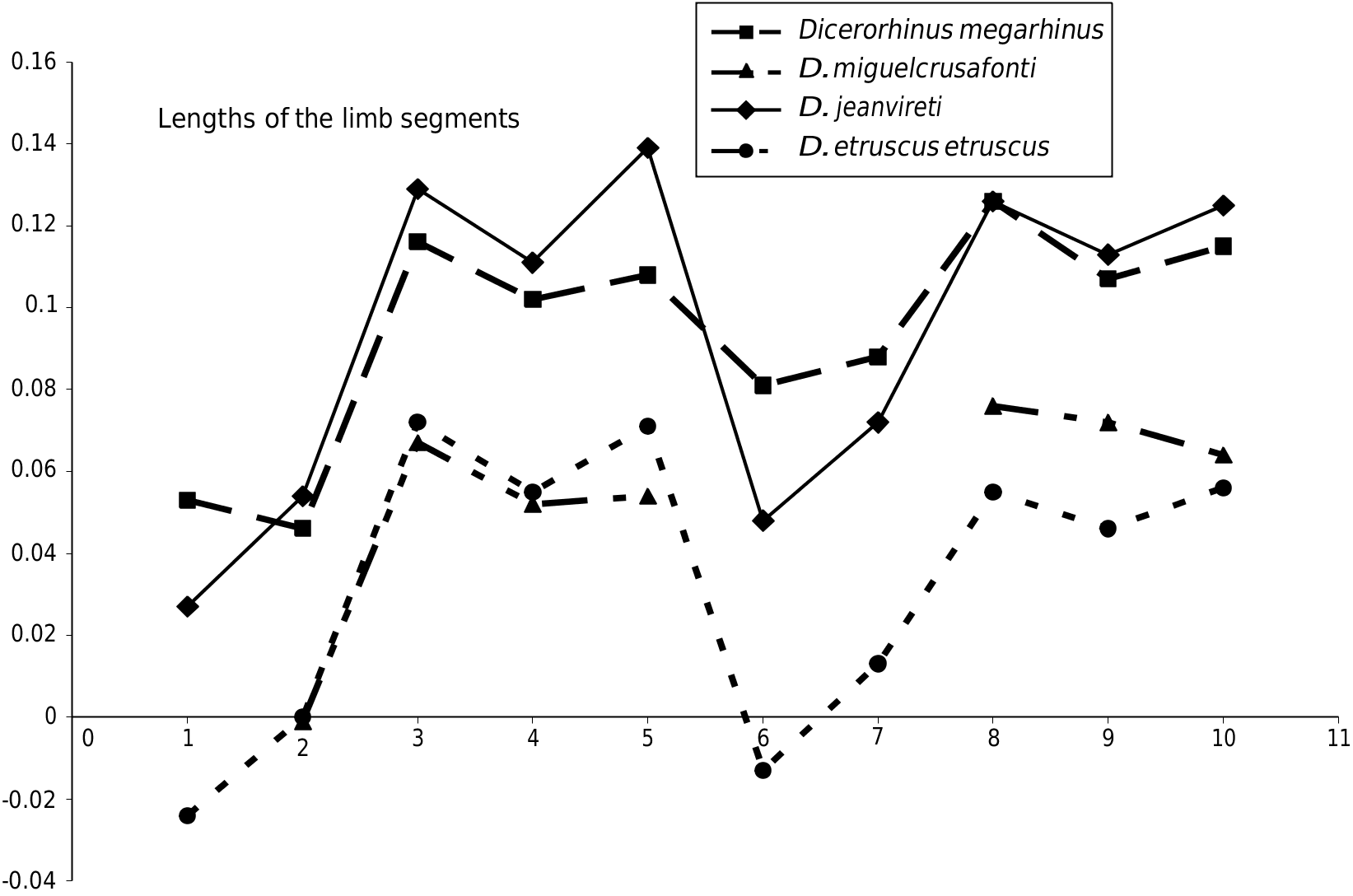
OF THE RHINOCEROS e Milia rhinoceros presents all the anatomical and biometric characteristics of D. jeanvireti . In Figure 15 View FIG , there is a comparative Simpson diagram of the lengths of the limbs for these four types of European rhinoceros considered in this article. It shows a certain similarity which reflects attribution to the same genus Dicerorhinus (the lengthening of metapodials, especially of abaxial ones, compared to Diceros bicornis , which serves as the reference material, is remarkable), and also shows that D. jeanvireti was more cursorial than D. megarhinus with its metapodials relatively longer, but shorter humerus, femur and tibia.
BIOSTRATIGRAPHIC IMPLICATIONS
D. jeanvireti is known from the zone MNQ 16, which corresponds to the Early Villafranchian. Considering the evolutionary trends highlighted in the European Dicerorhinus View in CoL by Guérin (1980), and for Rhinocerotidae View in CoL in general by Antoine (2002), it may be presumed that the Milia D. jeanvireti is a bit more advanced than that of Vialette; thus Milia could be a little more recent than Vialette, which lies at the base of the zone MNQ 16, but not younger than the Perrier-les Étouaires rhino, which is located near the top of the same zone. e age of Vialette is around 3 Ma and that of Perrier-les Étouaires is 2.5 + 0.1 Ma ( Guérin 2007).
PALAEOECOLOGICAL IMPLICATIONS e preferential habitat of Dicerorhinus jeanvireti is dominated by woodland with grassy areas that consist of graminae and ferns, in a humid climate ( Guérin 1980).
Order ARTIODACTYLA Owen, 1848 Sub-order SUIFORMES Jaeckel, 1911



| GSP |
Geological Survey of Portugal |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dicerorhinus jeanvireti Guérin, 1972
| Guérin, Claude & Tsoukala, Evangelia 2013 |
Stephanorhinus elatus
| RADULESCU C. 2005: 193 |
Stephanorhinus cf. elatus
| RADULESCU C. & SAMSON P. M. & PETCULESCU A. & TIUCA E. 2003: 3 |
S. elatus
| RADULESCU C. & SAMSON P. M. & PETCULESCU A. & TIUCA E. 2003: 3 |
Stephanorhinus jeanvireti
| LACOMBAT F. & MORS T. 2008: 160 |
| SPASSOV N. 2005: 203 |
| GLIOZZI E. & ABBAZZI L. & ARGENTI P. & AZZAROLI A. & CALOI L. & CAPASSO BARBATO L. & DI STEFANO G. & ESU D. & FICCARELLI G. & GIROTTI O. & KOTSAKIS T. & MASINI F. & MAZZA P. & MEZZABOTA C. & PALOMBO M. R. & NIO C. & ROOK L. & SALA B. & SARDELLA R. & ZANALDA E. & TORRE D. 1997: 171 |
| MAZO A. V. 1995: 287 |
| CAMPANINO F. & FORNO M. G. & MOTTURA A. & ZANO D. & SALA B. 1994: 452 |
Dicerorhinus jeanvireti Guérin, 1972: 133-136
| DURISOVA A. 2004: 99 |
| HOOIJER D. A. 1985: 35 |
| HOOIJER D. A. 1981: 472 |
| GUERIN C. 1972: 136 |
megarhinus
| HURZELER J. 1967: 636 |
Dicerorhinus megarhinus
| FEJFAR O. 1964: 89 |
megarhinus
| AZZAROLI A. 1963: 17 |
aff. megarhinus
| BOUT P. 1960: 188 |
etruscus
| DEPERET C. & MAYET L. & ROMAN F. 1923: 37 |
| FALCONER H. 1868: 367 |
elatus
| CROIZET J. B. & JOBERT A. 1828: 154 |