
Cryptophoca sp.
Peigné, Stéphane, 2016, Carnivora, Geodiversitas 38 (2), pp. 197-224 : 202-210
|
publication ID |
https://doi.org/ 10.5252/g2016n2a4 |
|
publication LSID |
urn:lsid:zoobank.org:pub:CDDFC6DE-E4D2-4001-9E8A-9B1CD6815B18 |
|
persistent identifier |
https://treatment.plazi.org/id/591C87F1-FFA4-3338-FBF9-EC52F5B0F9CE |
|
treatment provided by |
Felipe |
|
scientific name |
Cryptophoca sp. |
| status |
|
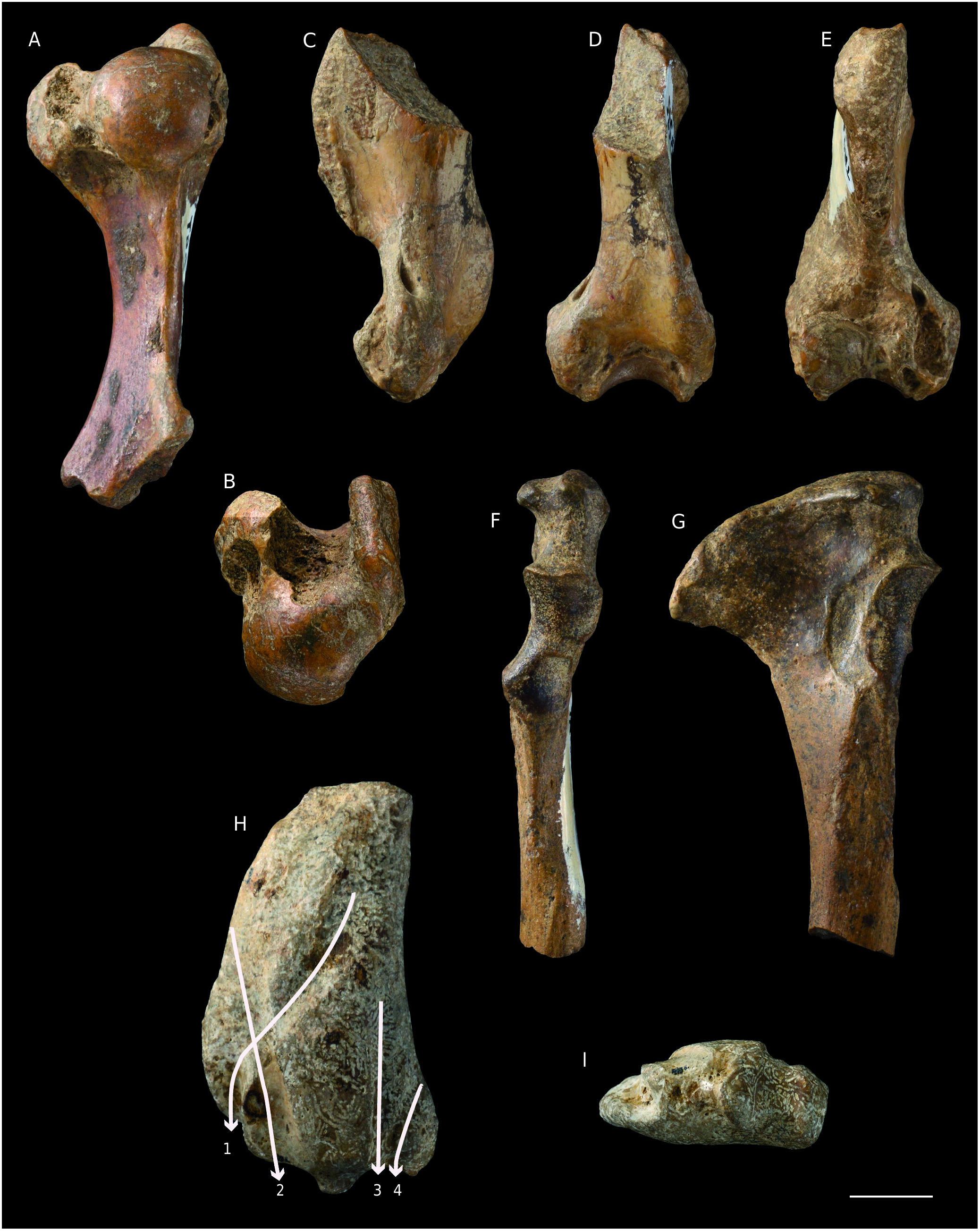
( Figs 1 View FIG F-H, 2, 3, 4A, B; Tables 2, 3, 4)
Phoca sp. – Malik & Nafiz 1933: 110, pl. 13, figs 1-3. Uncertain synonymy.
REFERRED MATERIAL FROM KÜÇÜKÇEKMECE. — KÇ 101, fragment of left hemimandible with roots of p2-4; MNHN.F.TRQ930, proximal two thirds of right humerus; TRQ933, proximal fragment of left humerus; TRQ935, distal half of right humerus; TRQ927 distal half of left radius; TRQ925, proximal half of left ulna; TRQ926, proximal fragment of right ulna; TRQ944, left femur without epiphysis; TRQ945 distal fragment of left femur without epiphysis; TRQ913, distal fragment of right tibia; TRQ916, distal fragment of left tibia; TRQ917, distal fragment of left tibia; TRQ918, distal fragment of left tibia; TRQ928, subcomplete left astragalus.
DESCRIPTION
Mandible ( Fig. 1 View FIG F-H; Table 2)
The dentary is broken off approximately one cm posterior to the distal alveolus of m1 and also lacks a small part in the anterior extremity (the canine alveolus is not completely preserved). The mandibular body is overall thick, but its ventral rim is pinched. Dentary depth reaches its maximum below p4-m1 (DMp4-m1 = 18.6), where the ventral rim of the body shows a marked convexity. This convexity has various names in the literature: digastric apophysis ( Grigorescu 1976), mental protuberance ( Koretsky & Ray 1994), chin prominence ( Koretsky & Grigorescu 2002). In the specimen from Küçükçekmece East, the mental protuberance is present though not marked. From this protuberance the dentary tapers off anteriorly and its ventral rim becomes oblique relative to the alveolar border. The symphysis is short and does not extend beyond the level of the mesial alveolus of p2; its surface is rugose only in its anterior part. On the labial face there are many mental foramina below p2 and partly coalescent ones below p3 and the diastema between p3 and p4. According to the preserved alveoli the p1 was large (nearly as large as the canine) and single-rooted, and the p2, p3, p4, m1 were biradiculate. Alveolar lengths indicate an increasing length from p2 to m1. The greatest diameter of the alveolus of p1 nearly reaches the p2 alveolar length. Diastemata are short, a few mm long, but they are present between all teeth; the longest diastema is the one between p4 and m1 (L = 4 mm). The tooth row (p1-m1, alveoli) is 48-mm long. The tooth row is not set strictly parallel to the long axis of the dentary, but it is slightly twisted, the p1 being set labially and the m1 being set more lingually.
Postcranium
Anatomical descriptions are based on the description of extant phocids ( Howell 1929; Piérard 1971) and middle-late Miocene taxa of Europe ( Koretsky 2001; Koretsky & Grigorescu 2002; Berta et al. 2015).
Humerus ( Fig. 2 View FIG A-E; Table 3). The humeri are robust and, though none is complete, they are complementary: MNHN.F.TRQ930andTRQ933 preserve the proximal part of this bone and TRQ935 preserves the distal two-thirds. The greater and lesser tubercles are greatly enlarged and projected far from the shaft. The greater and lesser tubercles extend proximal to the head. The humeral head is rounded, with mediolateral and anteroposterior (dorsoventral) diameters of similar size. The lesser tubercle is projected essentially to the medial and posterior sides. The intertubercular sulcus (or bicipital groove) is deep and has a maximum width of 7 mm. The deltopectoral crest is strongly developed and thick, especially at maximum crest height, where the deltoid tuberosity is prominent and slightly swollen laterally in TRQ933 and TRQ935; the crest is long and extends distally to approximately 10 mm from the distal articular surface. The epicondylar crest is prominent and anteroposteriorly thick, though not as developed and long as the deltopectoral crest. Nevertheless, it extends proximal to the distalmost point of the deltopectoral crest. The epicondylar crest projects mainly posteriorly and poorly laterad; the lateral rim of its anterior face is marked by attachments for the extensor muscles of the manus. It is not possible to compare directly the distal and the proximal epiphysis, butTRQ933 andTRQ935 appear to be adequately complementary, hence providing a good idea of the complete bone. The distal epiphysis is narrow, probably narrower than the proximal epiphysis, or of approximately equal width. The medial epicondyle is broad and anteroposteriorly flattened. An entepicondylar foramen is present. The olecranon fossa is nearly flat and the coronoid fossa is shallow. In anterior view, the capitatum is much wider than the trochlea.
Ulna ( Fig. 2 View FIG F-G; Table 3). The olecranon process is extremely expanded posteriorly and narrow in MNHN.F.TRQ925, much more so than in TRQ926, which probably belongs to a juvenile specimen. The olecranon crest is convex in lateral view. In anterior view, the proximal surface of the process is medially flared, with an anconeal process projected medially. The medial surface of the olecranon is depressed. The coronoid process of the trochlear notch is prominent, while the one of the radial notch is not. The trochlear notch is as tall as it is wide, while the radial notch is taller than wide. The shaft is broken off 3 cm distal to the radial notch; at this level it is reduced in diameter, oval in cross section.
Radius ( Fig. 2H, I View FIG ; Table 3). Only the distal articulation and a part of the shaft are preserved in MNHN.F.TRQ927. The shaft is strongly flattened mediolaterally and expanded anteroposteriorly. The anterior rim of the expanded region is convex, while the posterior rim is slightly concave. At least four grooves are present on the lateral surface for the tendons of extensor muscles of the manus. I interpret these grooves following the anatomical descriptions of Howell (1929) and Piérard (1971). The groove that obliquely crosses the distal region of the lateral side of the radius in a posteroproximal to anterodistal direction is particularly deep in the anterodistal corner. The position and the course of this groove indicate that this is the one for the tendon of the M. abductor pollicis longus (see Piérard 1971: fig. 20; Fig. 2H View FIG ). A wide, smooth groove located just posterior to it is interpreted as the groove for the M. extensor digitorum communis and/or that for the tendon of the M. extensor carpi radialis; these two tendons are very close one to another but are not always associated with a groove in extant phocids ( Piérard 1971). In the proximal part of TRQ927 this groove is visible along the anterior rim of the radius; distally it crosses the groove for the tendon of the M. abductor pollicis longus. A wide and low ridge separates this groove from two distinct grooves that are certainly for the tendons of the M. extensor digitorum lateralis and, more posteriorly, the M. extensor pollicis longus, as in extant phocids ( Howell 1929: fig. 10). A small facet for the scapholunar occupies the distal epiphysis; it is concavoconvex and, in distal view, figure-8 shaped. Distally, the posterior side is flattened and occupied by the facet for the ulna, which is elliptical and slightly depressed.
Femur ( Fig. 3 View FIG A-D; Table 4). The two specimens from Küçükçekmece West do not preserve the epiphysis and certainly belong to juvenile or young adult individuals. The absence of a deep trochanteric fossa in MNHN.F.TRQ944 is most probably a juvenile feature, hence has no pertinent taxonomic value. The lesser trochanter is absent. The greater trochanter is large, anteroposteriorly flattened and extended proximal to the femoral head. Its proximomedial part extends posteriorly above the trochanteric fossa (nearly absent/not yet developed? in TRQ944). The neck is short. The shaft is strongly anteroposteriorly flattened and has a lateral and, especially, a medial rim that is particularly thin at mid-length. The intercondyloid fossa is shallow. The main difference between TRQ944 and TRQ945 is that the latter shows a greater medial expansion of the medial epicondyle of the distal epiphysis. As TRQ945 belongs to a larger individual this difference may result from an ontogenetic difference. Indeed Koretsky (2001: 25) mentioned that juveniles tend to have limb bones where the proximal and distal epiphyses are not much different, while in adults the distal epiphysis is “distended considerably” compared to the proximal epiphysis.
Tibia ( Fig. 3 View FIG E-H; Table 4). The four distal extremities assigned to Cryptophoca sp. have nearly the same size and proportions,MNHN.F.TRQ917 being slightly larger. They all have similar morphology, except in the length of the preserved portion of the shaft. In its proximalmost part, the shaft is triangular in cross-section, then becomes more quadrangular distally. Distally on the medial face, there is a flattened, elongated triangular area corresponding to the surface of contact with the fibula. The medial side of the posterior face is marked distally by two grooves, interpreted here as those for the tendon of the M. flexor digitorum longus and, more medially, for the tendon of the M. tibialis posterior ( Piérard 1971: fig. 32). The grooves are well distinct in TRQ916 and TRQ913, but less so in TRQ917 and TRQ918, where only one groove is clearly identified. Lateral to these grooves, the tibia is convex and slightly bulged. The anterior face is marked by a wide groove, assigned to the one for the tendons of Mm. tibialis anterior and extensor hallucis longus. The anterior distal rim of the tibia extends distally to the level of the medial malleolus. The distal epiphysis is occupied almost entirely by the facet for the astragalus, which is strongly concave. Laterally the articular surface is inclined proximally, contrasting with the medial malleolus, which extends distally.
Astragalus ( Fig. 4A, B View FIG ; Table 4). The bone is poorly preserved. The calcaneal process is broken off and the articular facets are not well visible. The superior facet is proximodistally convex and mediolaterally flat. The neck is long and poorly deviated medially. The head, with an articular facet for the navicular, is flat distally and convex ventrally: the facet extends along the ventrolateral face of the neck. The facets for the calcaneus are not well distinct; the sustentaculum facet is narrow, convex and connected to the distal facet (for the navicular); the ectal facet is concave, wider, and separated from the sustentaculum facet by a shallow groove where a minute foramen is visible. The facet for the fibula occupies the entire lateral face of the body.
COMPARISONS AND DISCUSSION
Malik & Nafiz (1933: pl. 13, figs 1-3) figured and briefly described a craniodental material that was much more complete than that available today. Since their study, a fire destroyed all the specimens figured. The material found by Nicolas (1978) includes only post-cranial, mostly incomplete, elements.
Fossil phocids are abundant in late Miocene localities of Central Europe and the western and northern Black Sea region. In her review of the Miocene Phocinae Gray, 1821 of Europe Koretsky (2001: table 1) mentioned the species Monachopsis pontica (Eichwald, 1850) from Turkey, though no detail regarding the locality is mentioned. Among the many fossil Phocinae present in the middle and late Miocene of Europe, less than ten are known in eastern Europe, especially in the Black Sea region: Praepusa pannonica Kretzoi, 1941 , P. vindobonensis (Toula, 1897) , P. magyaricus Koretsky, 2003 , Sarmatonectes sintsovi Koretsky, 2001 , Histriophoca alekseevi Koretsky, 2001 , Monachopsis pontica and Cryptophoca maeotica . The taxonomic nomenclature follows Koretsky (2001). I also include in the comparisons Pontophoca sarmatica (Alekseev, 1924) , known from late Miocene strata of the same Black Sea region; Koretsky (2001) assigned this species to the Monachinae Gray, 1869 .
There is a great variability of body size among these late Miocene species. Cryptophoca sp. fromTurkey is a rather small species though a part of the material probably belongs to juveniles. Nevertheless I do not consider size as a pertinent character for taxonomic assignment.Pinnipeds generally show a marked sexual size (and morphological) dimorphism, but this is partly due to relative development of soft tissue (muscles, fat). Sexual dimorphism of bones does exist, and deals not only with size but also with the anatomy,which is worth knowing in the present context.In fossil seals, sexual dimorphism is generally not documented due to a lack of pertinent material. However, Koretsky (2001) provides differential measurements and traits for (supposed) males and females of Monachopsis pontica , which is one of the best-known late Miocene species. Praepusa vindobonensis also shows a great size variability ( Koretsky 2001). Below I therefore discuss chiefly anatomical differences rather than size differences.
Dentary
Morphologically the hemimandible from Küçükçekmece East closely resembles that of Cryptophoca maeotica illustrated by Koretsky & Ray (1994: fig. 1) and Koretsky (2001: fig. 33): same development and position of the mental protuberance, largesized and single-rooted p1, presence of diastemata between cheek teeth, development and length of the symphysis (straightness of the ventral rim, expansion not posterior to p2 mesial alveolus). The features that may represent autapomorphies for KÇ 101 and C. maeotica are the large size of the single-rooted p1 and the morphology of the anterior part of the dentary (symphysis length, ventral border straight).In these features these taxa differ from the species assigned to Praepusa Kretzoi,1941 , Histriophoca Gill, 1873 , Monachopsis Kretzoi, 1941 and Pontophoca Kretzoi, 1941 . KÇ 101 differs from C. maeotica in being smaller in size (by approximately 20%, based on illustrations of the tooth row of C. maeotica ; Koretsky & Ray1994:fig.1; Koretsky2001:fig.33) and in having an m1 longer than p4 (based on alveolar length). Postcranium
Comparison with C. maeotica . The humerus (MNHN.F.TRQ930, TRQ933, TRQ935) also fits the description of the same element in C. maeotica by Koretsky & Ray (1994) and Koretsky (2001): deltopectoral crest strong and long (though not reaching the coronoid fossa), lesser and greater tubercles at the same level and higher than the head, epicondylar crest reaching the distal part of the deltopectoral crest proximally.The fragmentary femora (TRQ944, TRQ945) belong to juveniles. They have a greater trochanter that is much higher than the head and a distal epiphysis that is only slightly wider than the proximal epiphysis, like in Cryptophoca maeotica ( Koretsky 2001: fig. 35).
Comparison with M. pontica . The description of the humerus from Küçükçekmece West also fits with that of the humerus of Monachopsis pontica . Given the sexual dimorphism observed in the latter ( Koretsky 2001) the distinction between the humeri of these species is not easy. According to the diagnosis of M. pontica provided by Koretsky, the humerus fragments from Turkey differ from the humerus of M. pontica in the absence of dorsoventral compression of the head and a shorter deltopectoral crest. However, the compression of the head in M. pontica is a highly variable, sexually dimorphic trait: the males have a dorsoventrally compressed humeral head, while the females do not ( Koretsky 2001: tables 2, 6). Koretsky (2001: fig. 17, table 7) describes the strong morphological and metrical sexual dimorphism of the femora of M. pontica . Juvenile characters may add to this dimorphism. The femora from Küçükçekmece West cannot readily be distinguished from those of M. pontica . A single astragalus of M. pontica has been described ( Grigorescu 1976: fig. 6). The description does not allow distinction from MNHN.F.TRQ928 but the figures show that the distal region of the astragalus of M. pontica (neck and head especially) are much more robust and wider relative to the body than in TRQ928.
Comparison with Praepusa spp. Compared to the material from Küçükçekmece West Praepusa spp. have a more slender humerus with a dorsoventrally compressed head, a sharper deltopectoral crest, a lesser tubercle that is less protruding (and elongated along the shaft axis) and a distal epiphysis that is wider relative to the proximal epiphysis ( Koretsky 2001: figs 24, 28; Koretsky 2003), an ulna with a shaft that is more mediolaterally compressed and an olecranon process that is much less developed anteroposteriorly ( Koretsky 2001: fig. 26), though P. magyaricus has an ulna with a similar development of the olecranon ( Koretsky 2003: fig. 2). Koretsky does not describe the radius and tibia of P. vindobonensis in detail, but their measurements and proportions do not differ markedly from the same elements from Küçükçekmece West ( Koretsky 2001: figs 27, 28, 31, tables 10, 12).
Comparison with P. sarmatica . Based on Koretsky & Grigorescu (2002) and Koretsky et al. (2014), the material from Küçükçekmece differs from Pontophoca sarmatica (and this is also true for P. jutlandica Koretsky, Rahmat & Peters, 2014
though not from eastern Europe) in having a single-rooted p1, a humerus with a longer deltopectoral crest, a more reduced epicondylar crest, a distal epiphysis (probably) narrower than the proximal epiphysis, and a femur with a higher greater trochanter and a distal epiphysis that is proportionally narrower, though exceeding the width of the proximal epiphysis.
The ulnae, radii, tibiae and astragali from Küçükçekmece West cannot be compared with all the late Miocene phocids of Europe due to lack of comparative data; only some of these bones may be compared to one or several taxa (see above the comparisons with Praepusa spp. and M. pontica ). They are provisionally assigned to the same species as the dentary, humeri and femora based on their size. Given the difficulty of generic assignment of fragmentary specimens, the material assigned here to Cryptophoca sp. may prove to represent more than one species.
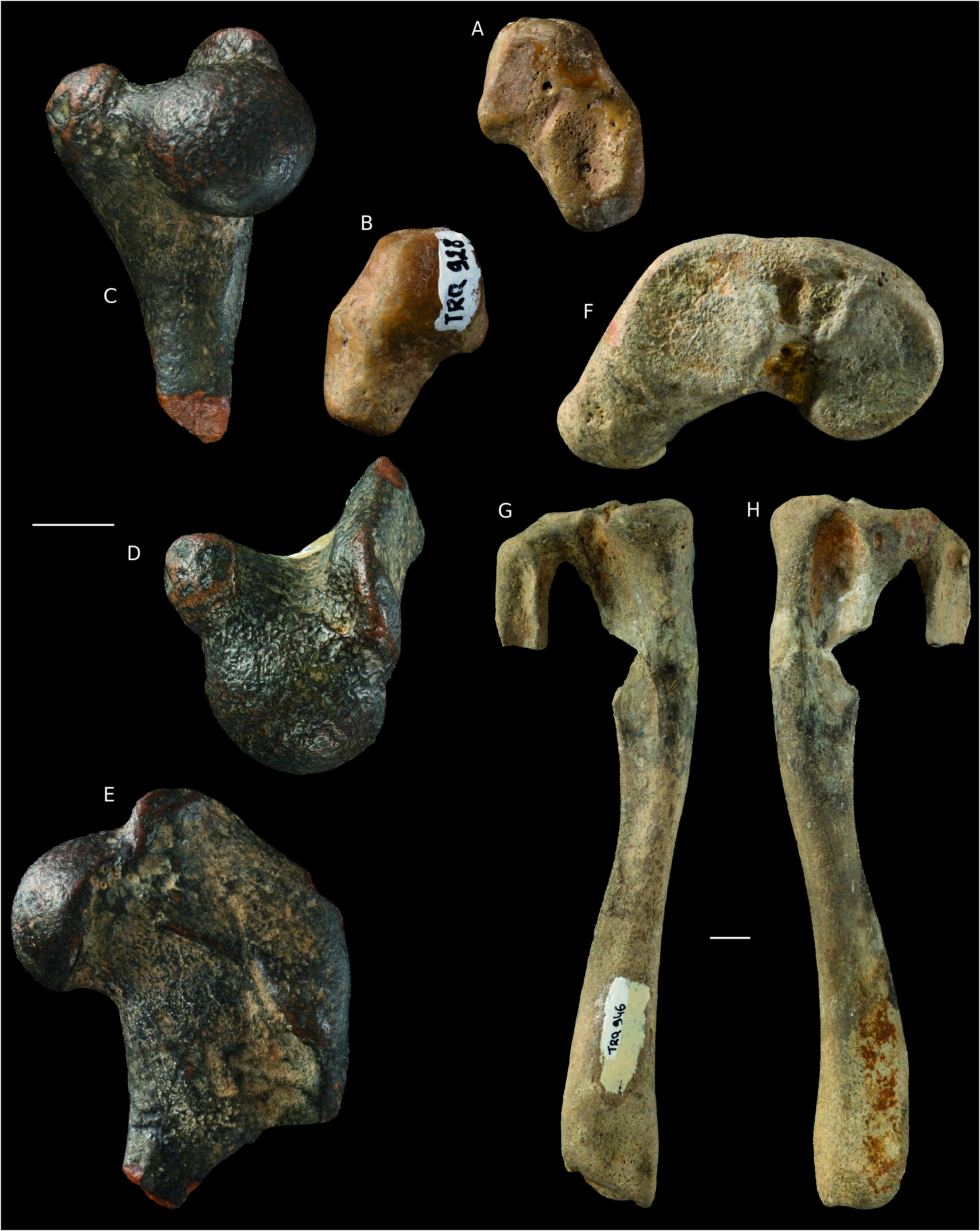
Phocidae gen. et sp. indet. ( Fig. 4 View FIG C-H; Table 5)
REFERRED MATERIAL FROM KÜÇÜKÇEKMECE. — MNHN.F.TRQ932, proximal fragment of right humerus;TRQ946, left tibia and proximal fragment of fibula; TRQ922, distal fragment of right tibia; TRQ923, proximal fragment of? left Mt I.
DESCRIPTION
Humerus ( Fig. 4C, D View FIG ; Table 5).
MNHN.F.TRQ932 belongs to a much larger specimen than those represented by Cryptophoca sp. The tubercles are developed and slightly higher than the humeral head. The latter is less rounded than in the specimens of Cryptophoca sp. described above, showing dorsoventral compression.
Tibia-fibula ( Fig. 4 View FIG F-H; Table 5).
The tibia and fibula are fused proximally (MNHN.F.TRQ946). Both articular facets for the femoral condyles are well defined and slightly concave. The lateral condyle is larger and more elliptical than the medial one. Just distally, and rather lateral, to the proximal epiphysis, there is a deep and short tibial fossa on the anterior and posterior faces. The anterior fossa is much shorter than the posterior fossa. The shaft is triangular in cross section; minimal diameters are measured slightly distal to mid-length (minimum mediolateral width is 14.6 mm, anteroposterior thickness is 13 mm at the same level). The distal part of the shaft and the distal epiphysis in TRQ922 and TRQ946 are not different from those of the specimens assigned to Cryptophoca sp. described above. The distal tip of the medial malleolus of TRQ922 is broken off. The grooves observed in the latter are visible in TRQ922, but in TRQ946 a single wide groove is distinct on the posterior face.
Approximately 30 mm of the distal fibula are preserved in TRQ946. The shaft is nearly T-shaped, with a posterior face divided by a sharp crest into two subequal parts. The anterior face of the shaft is slightly depressed by a fossa that may be the insertion area of the M. extensor digitalis lateralis, as in extant phocids ( Piérard 1971). Relative to the proximal face of the tibia, the proximal face of the fibula is inclined laterally distad.
Mt I ( Table 5) MNHN.F.TRQ923 may represent the proximal half of an Mt I. This identification is based on the morphology of the proximal epiphysis, which shows a strong concavity that may correspond to the articulation with the medial cuneiform; a small apophysis extending proximally is also present in the ventrolateral corner. Laterally, the facet for the Mt II is hardly visible.
COMPARISONS AND DISCUSSION
The fragmentary humerus MNHN.F.TRQ932 belongs to a much larger specimen than those represented by Cryptophoca sp. Besides size, its morphology differs from that of TRQ930 and 933 by having a deltoid tubercle located more proximally, a lesser tubercle oriented more medially, hence a much wider intertubercular sulcus, and a dorsoventrally compressed head. The latter character is also a characteristic of Monachopsis pontica and of some species of Praepusa ( Koretsky 2001) .
MNHN.F.TRQ946 and TRQ922 differ from those assigned to Cryptophoca sp. (TRQ913, TRQ916, TRQ917 and TRQ918) mainly by larger size (see Tables 4, 5). Fragmentary tibiae of Cryptophoca sp. are close in size to the tibiae of Praepusa vindobonensis , while TRQ922 and TRQ946 are of the size of Leptophoca lenis True, 1906 ( Koretsky 2001: table 12). In addition, compared to the specimens assigned to Cryptophoca sp. , TRQ922 and TRQ946 have a proportionally wider distal epiphysis.
I consider that the observed difference in size and anatomy between the humeri and tibiae of these two species cannot be attributed to sexual dimorphism and/or ontogeny.
Both the anatomical and taxonomic assignments of
MNHN.F.TRQ923 are judged to be provisional.
Suborder FELIFORMIA Kretzoi, 1945 View in CoL Family PERCROCUTIDAE Werdelin & Solounias, 1991


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.