
Chleuastochoerus linxiaensis, Hou, Sukuan & Deng, Tao, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3872.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:5DBE5727-CD34-4599-810B-C08DB71C8C7B |
|
DOI |
https://doi.org/10.5281/zenodo.6128155 |
|
persistent identifier |
https://treatment.plazi.org/id/B2017323-FFEE-FFA8-FF1D-FE1FDD0DC7CE |
|
treatment provided by |
Plazi |
|
scientific name |
Chleuastochoerus linxiaensis |
| status |
sp. nov. |
Chleuastochoerus linxiaensis sp. nov.
Figures 2–6
Holotype. IVPP V 18063 View Materials (Fig. 2a-d), cranium of a male individual (M3 almost completely erupted; the premaxillae, left zygomatic arch, pterygoid, tympanic bulla and occipital condyle are partly broken off; the canines, right P4-M3, left P1, P3-M3 are preserved).
Paratypes. IVPP V 18064.1-2 (Figs. 2e, 3, 4), skull of a juvenile female individual (M3/m3 erupting; left zygomatic arch, left pterygoid, and part of the occipital bone are broken off; right I1, C f, P1-M3, left I1-I2, C f, P1 and P4-M3 are preserved; the lower dentition is nearly complete, only the right i3 and c are broken). HMV 0 575 ( Fig. 5 View FIGURE 5 ), skull of a juvenile individual (M2/m2 erupting; zygomatic arch, pterygoid and occipital bone are partially broken off; left ascending ramus of the mandible is broken off; left and right I1-I3, C and P1-M1, left and right i1- i2, p1, p3-m1are preserved).
Localities and Horizon. Linxia Basin, Gansu Province, China. late Miocene, Liushu Formation, MN 10-13. V View Materials 18063 was collected from Yangwapuzi (LX0018, 35º24′49.5″ N, 103º30′28.6″ E) in Guanghe County, V18064.1- 2 from Hualinsanshe (LX0045, 35º23′36.7″ N, 103º25′46.9″ E) in Hezheng County, and HMV 0 575 from Shancheng (LX0041, 35º22′00.8″ N, 103º15′58.0″ E) in Hezheng County.
Etymology. The specific name linxiaensis refers to the Linxia Basin.
Diagnosis. Chleuastochoerus with longer facial region than in C. stehlini ; more anteromedial-posterolaterally compressed upper canine; strong pre- and postprotocrista, “protoconule” and “metaconule,” and lingual part of anterior and posterior cingula in P4; shorter trigonid, higher talonid, transversely arranged proto- and metaconid, and ridge-like ectoconid in p4; more pointed main cusps with deep and complex furrows of the cheek teeth; stronger anterior, posterior and buccal cingula in the upper molar.
Description. Pearson (1928) described the diagnostic characteristics of the genus Chleuastochoerus . Diagnosis of the new species is highlighted in the following paragraphs, and some amendments and supplements are proposed to Pearson’s description.
The cranium (Figs. 2–3, 5a–d; Table 1 View TABLE 1 ) of the new species is slender than in C. stehlini , and its dorsal profile has only a slight posterodorsal inclination. If the skull is divided into facial and cranial regions by the postorbital process of the frontal bone, the facial region is proportionally longer (ratios of facial/cranial regions is 2.52 in V 18064 View Materials and 2.57 in HMV 0575) than in C. stehlini (ratios: 1.86–2.40).
The snout of the male individual is broad at the supra-canine arch-niche; I1-I2 are arranged in a line about 45° to the median palatine suture, and I2-I3 about 60° to the suture; I3 is situated on the front wall of the supra-canine arch-niche. The snout in the female and juvenile individuals is narrow and the supra-canine arch-niche is weak, with I1-I3 about 30° to the median palatine suture. The supra-canine arch-niche is strong in the male individual but has a relatively thin and smooth rear wall which crosses obliquely with the facial region, whereas it is weak in female and juvenile individuals. On the dorsal wall of the supra-canine arch-niche there is a weak groove for the facial muscles to pass through.
The preorbital fossa is long, narrow and relatively shallow. A small and deeper depression can be recognized in the anteroinferior part of the preorbital fossa, which indicates the attachment area of M. dilatator naris lateralis. The facial region of the lachrymal bone is broad, and one small rounded lachrymal foramen is situated in front of the supra-anterior corner of the orbit, with its opening located postero-lateraly.
The palate is long, narrow and flat, with a U-shaped palate-maxillary suture that reaches the level between M1 and M2. The palatine foramen is situated in front of the palate-maxillary suture, near the level of the posterior part of M1. The choanae are broad and rounded, and the posterior border of the palate is situated 7 mm behind M 3 in IVPP V 18063 View Materials and separated from the pterygoid fossa by the thin pterygoid plate.
C. linxiaensis sp. nov. C. stehlini The ventral surface of the anterior part of the pre-zygomatic plate is depressed and separated by a ridge into two fossae, indicating the attachment area for the M. depressor rostri and M. masseter. The posterior part of the zygomatic arch is narrow and the ventral surface is crested for attachment of the M. masseter. The glenoid which is slightly convex longitudinally and weakly concave transversely, is slightly above the level of the basioccipital, and far above the level of the tooth row and the base of the paroccipital process; and there is a very weak post-glenoid process.
The tympanic bulla is relatively small and more laterally compressed than that of C. stehlini , situated close to the pterygoid fossa and projects antero-downwardly. The mastoid process is bifid, forming the lateral border of the relatively big stylomastoid foramen. The paroccipital process is thin and short, separated from the tympanic bulla by a narrow posterior lacerum foramen and projects nearly vertically. Two interconnected hypoglossal foramina appear inside the condyloid fossa.
The parietal bone is narrow, and the parietal crests are convergent significantly but do not formed a sagittal crest (the narrowest point is 9.6 mm in V 18063 View Materials ). The occipital bone extends far back and is divided by the external lateral crest of the occipital into a lateral part (temporal part) and a posterior part. The lateral part of the occipital surface, defined by the temporal crest and the external lateral crest of the occipital, is relatively small. The low and broad bell-shaped posterior part of the occipital surface hangs above the magnum foramen and mainly faces downward-backwardly. There is a weak crest extending from the external lateral crest of the occipital and ending at a weak tubercle. The end of the zygomatic arch, the mastoid process and the paroccipital process are imbricated to form a broad surface that is inferior to the posterior part of the occipital surface.
FIGURE 2. Cranium of Chleuastochoerus linxiaensis sp. nov. a-d, V 18063 View Materials (holotype), adult male cranium with C m, left P1, P4-M3, and right P4-M3; e, V 18064.1 (paratype), snout of juvenile female with I1, left I2, left and right C f and P1; a. dorsal view; b, ventral view; c, lateral view; d, occipital region view; and e, ventral view of snout. Scale bars equal 5 cm.
FIGURE 3. Juvenile female cranium (with right I1, C f, P1-M3, left I1-I2, C f, P1 and P4-M3) of Chleuastochoerus linxiaensis sp. nov. (V 18064.1). a, left lateral view; b, dorsal view; c, right lateral view; and d, ventral view. Scale bar equals 5 cm.
The parietal crests of the juvenile individual (HMV 0 575, Fig. 5 View FIGURE 5 a) are weakly convergent (the narrowest point is about 17 mm), and the temporal fossa is significantly short. There is a marked crest on the lateral side of the paroccipital process which extends up to connect with the external lateral crest of the occipital. Another weak crest starts from the external lateral crest of the occipital and ends at the tuber above the occipital condyle. The temporal crest is stronger than in the adult individual of both sexes. The pre-zygomatic plate is not completely formed. The preorbital fossa is shallow and the supra-canine arch-niche is weakly projecting. The snout is narrow as in the adult female individual. The diastemata between the incisors are relatively long.
The mandible ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 e–f; Table 2 View TABLE 2 ) is slender. The symphysis is fused in the juvenile individual, but the suture can still be seen. It is absent in adult individuals. The symphysis reaches the level of p2; it is notably broadened at the lower canine alveoli in the male individual, and is convergent at p1/p2. The lingual surface of the symphysis is smooth and deeply concave; whereas the mental surface is smooth from the lateral view, and slightly concave in ventral view, where there is usually a sagittal groove along the suture. The anterior border of the symphysis forms an angle of about 150º with the lower border of the horizontal ramus, and a pair of nutrient foramina is situated bellow i2 -c. The tooth rows are nearly parallel. The lower border of the horizontal ramus is nearly straight and only weakly curved at the angle of the mandible. The lateral surface of the horizontal ramus is relatively smooth, with a large mental foramen below the p1–p2 diastema. In the male individual, there is a rough bulge situated in front of a narrow depression on the lateral surface bellow p3–p4. The inferred position and the orientation of this bulge and depression are interpreted to be an anteroventrally descending unique branch of M. masseter, or a ventral branch of M. platysma ( Hou et al. 2014). The ascending ramus is relatively low and thin, with oblique anterior border and nearly vertical posterior border. The angle of the mandible is round and does not project backwards. The coronoid process is small and thin, the tip of which curves slightly backwards at nearly the same level or slightly lower than the mandibular condyle. The shape of the mandibular condyle is more or less cylindrical transversely, with a triangular plateau behind the articular surface.
The upper incisors (Figs. 3e, 5b; Table 3 View TABLE 3 ) are similar to those of C. stehlini , single-rooted, arranged in a broad arc in the male individual and narrow in the female individual. The I1 is the largest of the upper incisors; it is slightly compressed labial-lingually and forms an angle of about 45º angle to the sagittal plane; the postcrista is much longer than the precrista, and a small protuberance arises from the postcrista, which might be a budding cusplet; the enamel is smooth, only slightly swollen in the middle of the lingual side, and there is a weak lingual cingulum; there is a diastema between the I1 s, and the crown is usually worn from the tip to the postcrista. The I2 and I3 are notably small; the crown is more similar to the P1, which is more compressed and has the length greater than the width; the crown is semi-conical without wear, and the postcrista is longer than the precrista, which forms a line angled about 60º to the sagittal plane; the labial side of the crown is convex, and the lingual side is relatively flat; no cusplet or cingulum can be observed. The DI 1 is similar to the I1 but is relatively smaller and sharper; the ridge in the middle of the lingual side is very weak and there is no lingual cingulum; the long axis forms a smaller angle with respect to the sagittal plane. The DI 2 is also similar to the I2 but much smaller, and the crown is notably low; no cingulum or cusplet can be observed. The DI 3 is very simple but larger than the DI 2.
C. linxiaensis sp. nov. C. stehlini
*V18064.2 *HMV 0 575 V 18065 View Materials V 18066 View Materials V 18067 View Materials HMV 1439 * juvenile individual.
L, medial-distal length; W, labial-lingual width; *, deciduous teeth; l, left; r, right.
L, mesio-distal length; W, labial-lingual width; Li, lingual width; La, labial width; Po, posterior width; *, deciduous teeth; l, left; r, right.
The lower incisors ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 f; Table 4 View TABLE 4 ) are tightly arranged in a relatively broad arch in the male individual and narrower in the female individual. The i1 is high, with very strong ridge in the middle of the lingual side; its labial side is smooth and rounded. There is a short diastema between the roots of the i1 s, but their crowns contact each other. The i2 is more robust and higher than the i1; there is a strong central ridge in the lingual side, which is slightly distally located; the labial side is convex. The i3 differs from the i1 and i2; the crown is much smaller and sharper, with the height only slightly greater than the width; the pre- and the postcristids are strong, and the postcristid is longer than the precristid; the lingual side is smooth and slightly depressed with a clearly central ridge, and the labial side is smooth and slightly convex. There is a short diastema between the i2 and i3. The di1 and di2 are similar to the i1 and i2, but are much lower, smaller and more slender.
The upper canine (Figs. 2, 3, 6; Table 3 View TABLE 3 ) of Chleuastochoerus is specialized. This short and robust tooth is oriented mainly downwards and forwards and slightly outwards, but never upwards. The C m is like a pyramid that been anteromedial-posterolabially compressed; the antero-medial surface is oriented at about 45˚ to the tooth row plane and is slightly depressed near the root, with very thin enamel; the postero-labial surface is more convex but often has a medial depression; at the base of the contact between the antero-medial and the postero-labial surfaces there is a notable oval swelling which is covered by relatively thick enamel; a big and nearly vertical wear facet appears at the lateral part of the antero-labial surface of the C m. The C f is smaller and less compressed than the C m, and its cross section is like a compressed half arch; the medial side of the C f is nearly flat and rough without enamel; and the lateral side is slightly convex and covered by very thin enamel; the wear facet is on the anterior side, vertical but narrower than in the C m.
The C f ( Fig. 4 View FIGURE 4 b; Table 4 View TABLE 4 ) is slender and has a verrucosic cross-section, and is nearly vertical but curves slightly backwards. The enamel is very thin on the lateral and the lingual surface, and a wear facet covers most of the posterior side.
The paracone of the P1-P3 ( Figs. 6 View FIGURE 6 a, c; Table 5) is pointed, more compressed in the P1 and P2, and more conical in the P3, with sharp pre- and postcrista; in side view the outline is triangular. The postcrista is longer than the precrista, and there is a weak accessory cusp on the postcrista of the P3 ( V 18063 View Materials ). The protocone is a tiny pointed elevation on the talon of the P1, and enlarged in the P2 and P3, so the width of the posterior lobe of the P1 is similar to the anterior lobe, while the posterior lobe of the P2 and P3 is wider than the anterior lobe. There is a weak anterior cingulum which extends onto the anterior part of the lingual side in the P3, and the posterior cingulum is continuous and fused with the talon; there is no buccal cingulum. There is a short C-P1 diastema (3.4 mm) and a notably longer P1-P2 diastema (16 mm) in V 18063 View Materials . The P4 ( Figs. 6 View FIGURE 6 a, c; Table 5) is like a square or a rounded triangle in occlusal view, and its buccal side is slightly longer than the lingual side; the paracone and the metacone are well separated, with a groove present on the lingual and the occlusal sides, and partly visible at the top half of the buccal side; the paracone and the metacone are similar in size, and the paracone is slightly higher than the metacone; furrows can be observed on the lingual side of the paracone and the metacone, but there are no sagittal cusplets ( Pickford 1988); the protocone is similar in size to the paracone, and is situated slightly anterior to the midline of the paracone and the metacone, so the anterior border of the P4 is slightly inclined posteriorly; the pre- and the postprotocrista are short and steep, and the furrow is conspicuous; the preprotocrista contacts the low and small but isolated “paraconule,” and the postprotocrista contacts the low and small “metaconule”; the “metaconule” also contacts the posterior cingulum instead of the metacone, but does not fuse with the posterior cingulum; the anterior and posterior cingula are thick and continuous, and the buccal and lingual cingula are absent. The DP2 ( Figs. 5 View FIGURE 5 b–c; Table 5) is similar to the P2 but smaller and sharper with thinner enamel and a smaller talon. The DP3 ( Figs. 5 View FIGURE 5 b–c; Table 5) is nearly triangular in occlusal view, its crown is very low and the enamel is thin; the three main cusps are similar in size, and the anterior cusp is located on the extension of the midline between the two posterior cusps; there is no cingulum. The DP4 ( Figs. 5 View FIGURE 5 b–c; Table 5) is molarized; it has four main cusps, and a small medial accessory cusp which contacts the posterior main cusps when worn; the anterior and posterior cingula are thin, and the buccal cingulum is only visible at the posterior part of the teeth and is continuous with the posterior cingulum.
The p1–p3 ( Fig. 6 View FIGURE 6 b; Table 6) are relatively hypsodont with sharp protoconid and thin pre- and postcristids; the precristid is notably shorter than the postcristid in p1, and only slightly shorter than the postcristid in the p2 and p3; there is no accessory cusp in the p1, whereas the postcristid is weakly serrated in lateral view in p2 and p3; the talonid is very small, does not inflate or form a conid, so the crown width does not enlarge backwards. The p4 ( Fig. 6 View FIGURE 6 b; Table 6) is similar to p 3 in length, but is much wider than p3 by having a well-developed metaconid and a wellseparated talonid with a large cusp. The anterior border of the crown is steep in lateral view; the metaconid and the protoconid are nearly transversely situated and the protoconid is placed slightly forward; the protoconid is narrow and small, with a low and short preprotocristid which is inclined lingually and separated from the metaconid by a deep groove; the metaconid is more conical than the protoconid, bearing clear furrows on the postero-buccal side; the talonid is high and broad, separated from the trigonid by a deep and narrow valley; the ectoconid is crestshaped, transversely situated at the lingual-posterior corner of the metaconid; the anterior and the posterior cingula are thick and continuous, and the lingual and the buccal cingula are absent.
P1 P2 P3 P4 L×W L/W L×W L/W L×W L/W L×W L/W
C. linxiaensis sp. nov.
V 18063 View Materials l 8.4×3.7 2.27 9.3×7.2 1.29 9.0×10.5 0.86
r 9.2×10.0 0.92
V 18064.1 l 8.4×3.8 2.21 10.3×10.1 1.02 r 8.3×3.8 2.18 *8.1×3.7 *2.19 *9.6×6.8 *1.41 *10.1×9.0 *1.12
L, antero-posterior length; W, labial-lingual width; *, deciduous teeth; l, left; r, right.
The M1 and M2 ( Figs. 6 View FIGURE 6 a, c; Table 7) are similar, but M2 is notably larger than the M1. The M1 is nearly rectangular with its length slightly greater than the width, and the posterior lobe is slightly broader than the anterior lobe; whereas the length of M2 is much greater than its width, and the anterior lobe is wider than the posterior lobe. The main cusps are similar in size and more pointed than in C. stehlini ; the furrows of the main cusps are deep and long, almost reaching the top of the cusps, and there are sub-furrows which make the main cusps more complex; the anterior accessory cusp (= protoconule of van der Made 2010) is triangular, connected to the protocone and separated from the top of the anterior cingulum by a weak groove in V 18063 View Materials ; the medial accessory cusp is big and high, bearing weak furrows; the posterior accessory cusp is small and low, located between the metacone and the posterior-medial furrow of the hypocone; the anterior and the posterior cingula are thick and continuous; the buccal cingulum can be only observed on the buccal valley, but is high and continuous in V 18063 View Materials which is suspected to be atypical; the lingual cingulum is only weakly seen in the medial valley. The M3 ( Fig. 6 View FIGURE 6 c; Table 7) is nearly triangular due to its notably small talon; the main cusps of the M3 are similar to those of the M2, but the furrows are less complicated than in the M2; the accessory cusps are larger and are more isolated compared with the M2, but the posterior accessory cusp is small and bifid; the anterior and the posterior cingula are thick and continuous, and the buccal and lingual cingula are only weakly seen in the buccal and the medial valley.
p1 p2 p3 p4 L×W L/W L×W L/W L×W L/W L×W L/W
C. linxiaensis sp. nov.
V 18064.2 l 7.3×3.1 2.36 8.9×3.2 2.78 10.0×4.4 2.27 11.6×7.4 1.57 L, antero-posterior length; W, labial-lingual width; *, deciduous teeth; l, left; r, right.
The m1-m2 ( Fig. 6 View FIGURE 6 b; Table 8) are rectangular, with the m2 being larger than m1. As in the upper molars, the furrows of the main cusps are complex; the anterior accessory cups (we prefer to call it “preprotocristid”) are small, and it is separated from the anterior cingulum by a narrow groove; the medial accessory cusp is notably large, connected with the hypoconid and the ectoconid at the base; the posterior accessory cusp is big and strong, and swells anteriorly from the posterior cingulum; the anterior and the posterior cingula are thick and continuous, and the buccal cingulum only weakly seen in the buccal valley of HMV 0 575 and absent in V 18064.2. Only V 18064.2 has an erupting m3 ( Fig. 6 View FIGURE 6 b); the protoconid and the metaconid are notably sharp, bear complicated furrows and sub-furrows; the anterior accessory cusp is small and thin, weakly bifid and fused with the anterior cingulum; the medial accessory cusp is smaller than in m2, and is bifid.
Comparisons. Chleuastochoerus was described to include Schlosser’s “ Sus stehlini ”. Unfortunately, neither Schlosser (1903) nor Pearson (1928) designated a holotype for C. stehlini . “ S. microdon ” was also described by Schlosser (1903) for two isolated third molars which are smaller than those of “ S. stehlini ”. Pearson (1928) disagreed that “ S. microdon ” was a small variety of Chleuastochoerus because the age and locality of the material is uncertain and also because very similar suid teeth may belong to very different types of skulls. However, van der Made & Han (1994) evaluated the size differences, and suggested that these differences reflected evolution and time, not geographical or ecological differences between contemporaneous populations. An evolutionary transition from “ S. microdon ” to “ S. stehlini ” was proposed by van der Made & Han 1994 and van der Made 2010; the materials from Loc. 49 represented “ S. stehlini ”, and other materials from Loc. 12, 29, 71, 73 represented “ S. microdon ” (van der Made 2010). Since Schlosser’s type material was unavailable to van der Made, the hypothesis was proposed but no new species was erected.
C. linxiaensis sp. nov.
V 18063 View Materials l 12.9×11.8 1.09 16.3×13.3 1.23 ca.17.0×13.4 1.26 r 13.6×11.8 1.15 17.0×13.5 1.26 ca.18.0×14.0 2.07
V 18064.1 l 14.0×11.7 1.20 17.6×13.0 1.35
r 13.7×11.9 1.15 17.6×13.6 1.29
HMV 0 575 l 12.1×11.7 1.03
r 13.6×11.5 1.18
V 18061 View Materials l 12.9×12.1 1.07 15.6×13.2 1.18 21.4×13.9 1.54 r 13.2×11.9 1.11 16.0×13.0 1.23 21.6×13.7 1.58
V 18062 View Materials l 11.8×11.9 0.99 14.4×13.3 1.08 22.2×12.9 1.72 r 11.7×11.9 0.98 14.3×13.0 1.10 20.0×13.2 1.52
HMV 0 569 l 13.4×11.9 1.13 15.9×13.6 1.17 20.0×14.0 1.43 r 13.3×12.0 1.11 16.0×13.7 1.17 20.2×13.9 1.45
HMV 1439 l 11.9×11.2 1.06 14.4×12.0 1.20 19.6×12.1 1.62 r 12.0×11.0 1.09 14.3×12.2 1.17 19.8×12.2 1.62
HMV 1870 l 14.5×12.7 1.14 15.6×13.3 1.17 17.8×13.8 1.29 r 14.3×12.6 1.13 15.8×13.6 1.16 17.3×13.6 1.27
HMV 0 156 l 12.6×10.8 1.17
r 12.6×11.5 1.10
L, antero-posterior length; W, labial-lingual width; l, left; r, right.
The M3 of “ Sus microdon ” falls within the size variation of the materials from Loc. 49 and the Linxia Basin, and its morphology is also similar to the M3 of Chleuastochoerus ; whereas the m3 is shorter than all the known m3 of specimens of Chleuastochoerus . Since the age and localities of Schlosser’s materials are uncertain, the m 3 may belong to another small suid form. Van der Made used the anterior width of the m3 to show the size variation in the Chinese Chleuastochoerus (see van de Made 2010: p. 108, fig. 35), and the data provided by Pearson (1928) did show a size variation between the materials from Loc. 49 and from other localities. However, when we add the data of the materials from the Linxia Basin, a continuous increase is revealed by the materials from different localities ( Figs. 7 View FIGURE 7 , 8 View FIGURE 8 ), thus it is hard to define a different species on the basis of size. Certainly the width increase is more notable than the length, but one thing to note is that the molar length in ungulates is claimed to be much less affected by diet than its width ( Damuth 1990; Fortelius 1990).
As mentioned above, two morphotypes of Chleuastochoerus are found in the Linxia Basin. The first morphotype has a relatively short facial region, and more globular main cusps and relatively simple furrows in the cheek teeth (Figs. 9, 10). These features resemble the cranium from Loc. 49 ( Pearson 1928, text-figs. 6–9, now housed at the Museum of Paleontology, University of Uppsala, Sweden, with specimen Number M 3679) which was described by Pearson in detail. Since the syntype of C. stehlini was perhaps lost during the Second World War, the cranium from Loc. 49 (M 3679) is designated as the neotype of C. stehlini , and the materials of the first morphotype from the Linxia Basin are assigned to C. stehlini .
L×W L/W L×W L/W L×W L/W
C. linxiaensis sp. nov.
V 18064.2 l 17.0×11.3 1.50
r 12.0×9.5 1.26 17.1×11.2 1.53
HMV 0 575 l 13.0×
r 12.8×9.3 1.38
V 18065 View Materials l 12.0×9.1 1.32 14.4×10.3 1.40 22.5×10.8 2.08 r 12.3×9.0 1.37 14.5×10.2 1.42 22.7×10.9 2.08
V 18066 View Materials l 12.5×9.3 1.34 15.0×10.6 1.42 22.8×10.6 2.15 r 12.7×9.3 1.37 22.5×11.1 2.03
V 18067 View Materials l 12.0× 15.6×11.0 1.42 24.3×11.2 2.17 r 12.9×9.1 1.42 15.8×11.2 1.41 24.0×11.7 2.05
V 18068 View Materials l 11.3×8.2 1.38 14.1×9.7 1.45 20.9×10.6 1.97 r 11.8× 13.6×9.8 1.39 20.6×10.5 1.96
V 18069 View Materials l 12.1×8.6 1.41 14.3×10.3 1.39 21.0×10.9 1.93 r 12.0×8.4 1.43 13.8×10.4 1.33 21.0×11.0 1.91
HMV 0 564 l 12.9×8.9 1.45 15.7×10.5 1.50 23.4×11.1 2.11
r 13.1×8.8 1.49 15.6×10.6 1.47
HMV 0 565 l 12.4×8.5 1.46 14.6×10.4 1.40 20.5×10.7 1.92 r 12.5×8.5 1.47 14.6×10.2 1.43 20.2×10.7 1.89
HMV 0 566 l 14.2×9.2 1.54 15.5×11.2 1.38 26.1×12.1 2.16 r 25.5×12.0 2.13
HMV 1439 l 12.0×9.6 1.25 14.4×10.5 1.37 22.2×11.3 1.96 r 12.2×8.8 1.39 14.9×10.4 1.43 22.4×11.3 1.98
L, antero-posterior length; W, labial-lingual width; l, left; r, right.
The second morphotype bears relatively long facial region and more complex cheek teeth, this second morphotype is described in this report as Chleuastochoerus linxiaensis sp. nov. Comparison of the materials described by Pearson (1928) and the C. stehlini materials from the Linxia Basin shows that the new species differs from C. stehlini in the following features (the characters of C. stehlini are listed between brackets): the dorsal profile of the cranium is only slightly inclined postero-dorsally (more inclined); the parietal crests converge more rapidly (weakly in the young individuals, and moderately in the old individuals); the facial region is relatively long and narrow (shorter and stouter), in the juvenile individual (HMV 0575), the ratio of 100×facial region/cranial region is extremely large (256.9); the rear wall of the supra-canine arch-niche is thin and smooth (thick and rough, usually with exostosis); the paracone and metacone of the P4 are nearly equal in size and are clearly separated, and the pre- and postprotocrista, the “protoconule” and “metaconule” are strong (the metacone is usually smaller than the paracone and is weakly separated from the paracone, the cristae and accessory cusps of the protocone are usually weak or absent); the meta- and the protoconid of the p4 are nearly transversely placed and well separated, and the preprotocrista is short and projects antero-lingually, so that the anterior margin is nearly vertical in side view and the trigonid is broad and short (the preprotocrista projects more anteriorly so that it forms a “paraconid” in some specimens, the trigonid is long); the talonid of the p4 is large and high, separated from the trigonid by a deep groove, and has a ridge-like ectoconid situated postero-lingually to the hypoconid (the talonid is short and low, only a ridge-like hypoconid is longitudinally located, and there is no ectoconid); main cusps of the cheek teeth are more pointed and bear more complicated accessory cusps and furrows (more globular main cusps with simpler accessory cusps and furrows); the anterior and posterior cingula of the upper molars are thicker, and the lingual and the buccal cingula sometimes present in the upper molars (only weak anterior and posterior cingula).
Vislobokova (2009b) described Chleuastochoerus tuvensis , from Russian specimens, and listed six features that separate C. tuvensis from C. stehlini : 1) short facial region; 2) larger I3 (alveoli not reduced); 3) P4 with widely separated paracone and protocone, and the protocone displaced anteriorly; 4) the M3 narrowing posteriorly and having a weaker buccal cingulum; 5) weaker narrowing of the lower jaw at the p1–p2 diastema; 6) the absence of a p2–p3 diastema. Based on our observation and comparisons, characters 2, 5, 6 may be individual or sexual differences, and the short facial region is also doubtful for it is inferred from the badly compressed mandible. Nonetheless, characters 3 and 4 do differentiate C. tuvensis from C. stehlini . Besides, the P4 of C. tuvensis has a larger accessory cusp than in C. stehlini , and the anterior cingulum and the accessory cusps of the M 3 in C. tuvensis are also stronger than in C. stehlini . Moreover, in C. tuvensis , the main cusp of the p3 is more anteriorly displaced with steep anterior side and the talonid of the p3 and p4 is higher and longer; the buccal valley of the molars is deeper, and the accessory cusps of the m3 are larger than in C. stehlini . All these prove the validity of C. tuvensis , which preserves more progressive cheek teeth than C. stehlini . The new species differs from C. tuvensis in the following aspects: the P4 of C. tuvensis is larger than that of the new species; and the paracone and protocone of the P4 are also more widely separated in C. tuvensis than in the new species; the P4 of C. tuvensis has a big accessory cusp behind the protocone, so the protocone is notably anteriorly located, whereas, the P4 of the new species does not have accessory cusp, but bears notable pre- and postprotocrista and “protoconule” and “metaconule”; the crown of the upper molars in C. tuvensis is narrower and its buccal valley is deeper than in the new species, and the anterior and the posterior cingula are thicker and higher in C. tuvensis than in the new species; unlike the new species, C. tuvensis does not have a buccal cingulum; the protoconid of the p 4 in C. tuvensis is anteriorly situated with a small “paraconid” placed antero-buccally to the metaconid, which resembles C. stehlini and differs from the new species; and the p4 of C. tuvensis has high and big talonid like in the new species, but lacks an ectoconid.


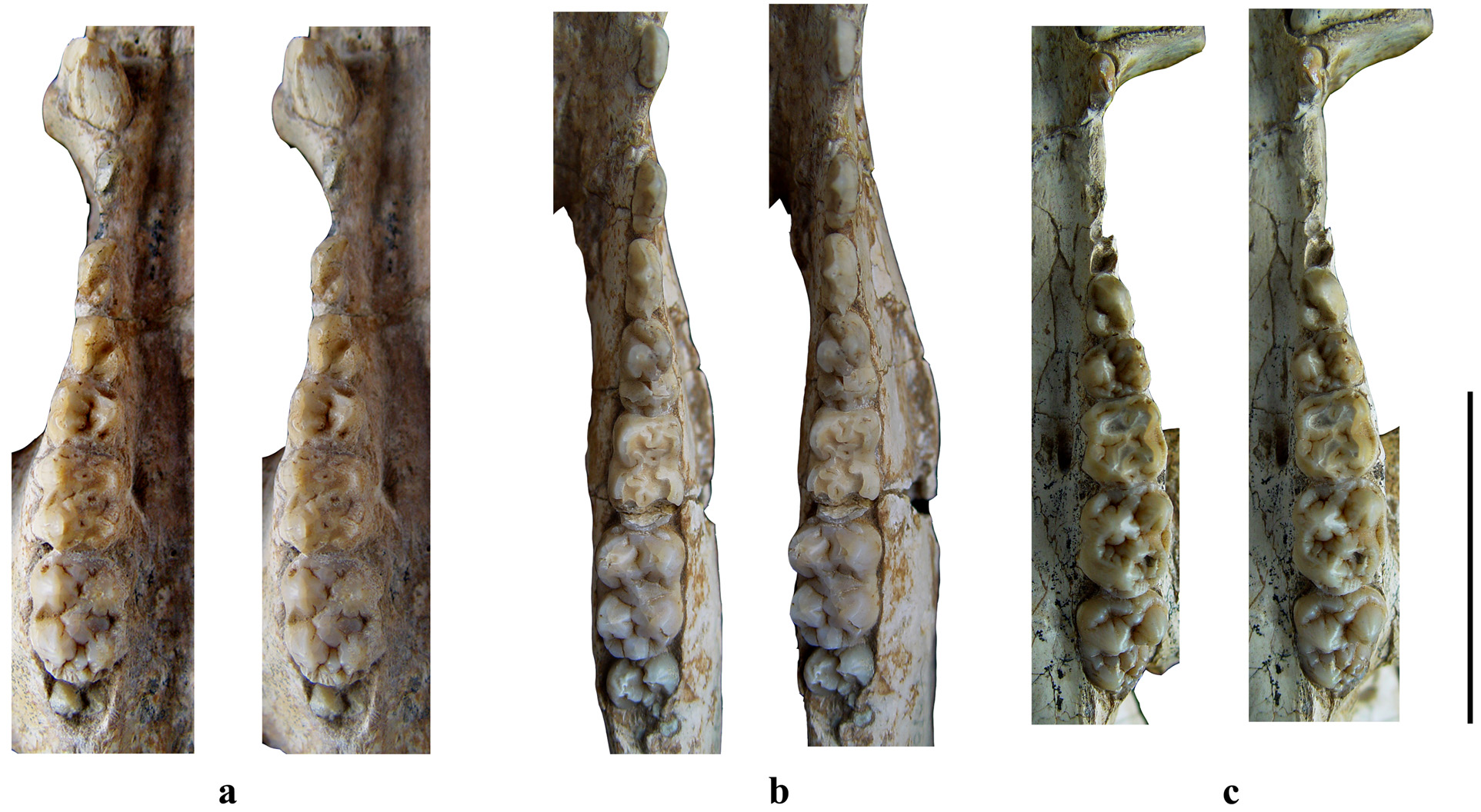
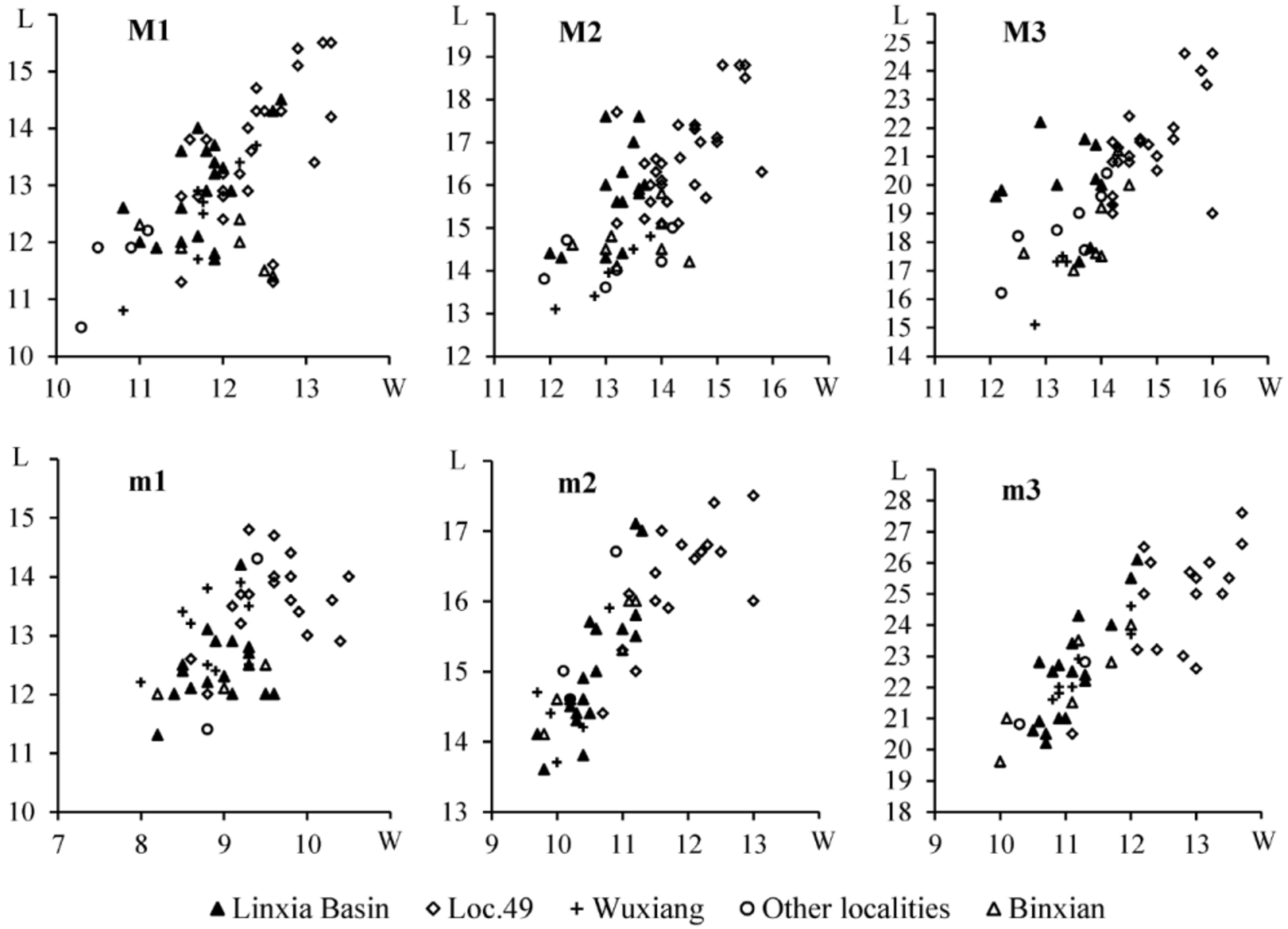
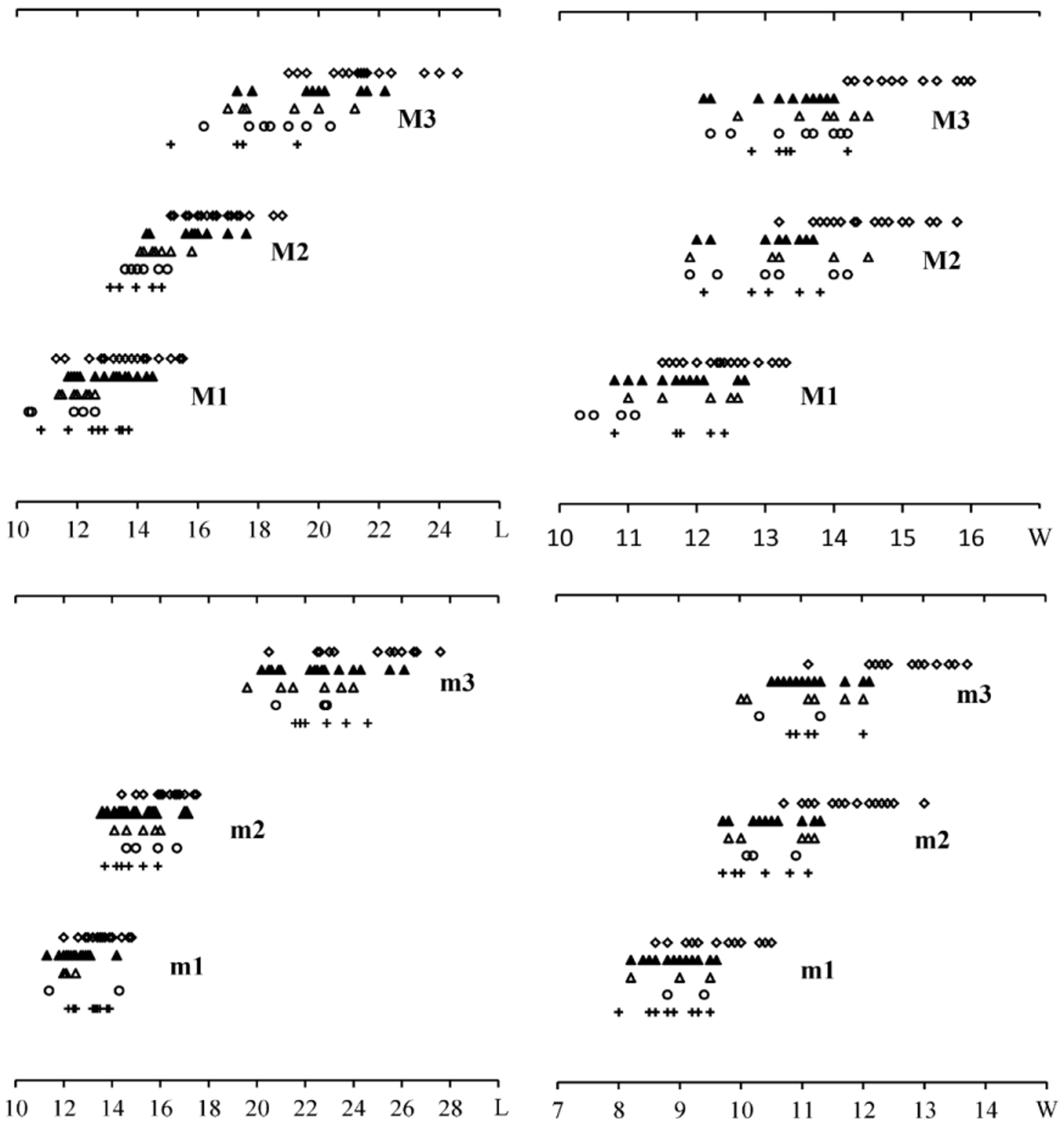
TABLE 1. Cranial measurements (in mm) of Chleuastochoerus from the Linxia Basin.
| V 18063 View Materials *V 18064.1 | *HMV 0 575 | V 18061 View Materials | V18062 View Materials HMV0569 | HMV 1439 | *HMV 0 156 | |
|---|---|---|---|---|---|---|
| 1. Maximal length of cranium | >241.0 | 202.2 | 250.7 | 268.4 257.3 | 268 | 214.1 |
| 2. Basal length of cranium | 212.0 | 182.4 | 203.9 | 216.5 227.4 | ca. 215.5 | |
| 3. Width behind paroccipital process | 69.5 ca.65.0 | 68.0 | 83.1 | 76 78 | >67 | 56.8 |
| 4. Minimal width between parietal crests | 9.6 ca.13.0 | 16.6 | 12.5 | 9.1 27.3 | 12.4 | 16.9 |
| 5. Length of facial region | 181.5 | 149.0 | 166 | 183.7 191 | 189.5 | 151.8 |
| 6. Length of cranial region | 79.0 ca.72.0 | 58.0 | 89.2 | 93.6 79.6 | 87 | 69.4 |
| 7. Height of occipital region | ca.67>67.5 | 59.4 | 84.5 | 81.6 74.4 | ca.83.5 | |
| 8. Height above the foramen magnum | 51.0>46.0 | 41.0 | ca. 65 | 61.4 56 | ||
| 9. Width of orbit | 27.8 28.6 | 24.2 | 25 | 32 29.5 | ca.27 | 27.0 |
| 10. Height of orbit | 28.5 27.9 | 28.0 | 40.6 | 33.5 38.5 | ca.40 | |
| 11. Width at supra-canine arch- niche | 68.5 ca.46.0 | 86.4 | ca. 85 ca. 70 | 47.5 | ca.35.0 | |
| 12. Premaxillae – posterior border of palate | 153.4 | 126.7 | ?151 | 155?161 | 157.5 | |
| 13. M3 - posterior border of palate | ca.7.0 | ?13.4 | 17.3?20 | 23.4 | ||
| 14. P1-M3 | ca.95.0 | 87 | 85.8 ca. 90 | 86.4 | ||
| 15. P1-P4 | ca.50.0 43.2 | 40.8 | 39.4 | 40.2 ca. 41 | 41.3 | 38.5 |
| 16. M1-M3 | ca.45.0 | 48.5 | 46.4 49.2 | 45.5 | ||
| ratios/% | ||||||
| 1) 3:1 2) 5:6 | 27.0 252.1 | 33.6 256.9 | 33.1 186.1 | 28.3 30.3 192.3 239.9 | 25.0 217.8 | 26.5 218.7 |
| * juvenile individual. |
TABLE 2. Mandibular measurements (in mm) of Chleuastochoerus from the Linxia Basin.
| 1. Mandible length (without incisors) 177.8 | 146.3 | 182.9 | 187.3 | 176.9 | 181.2 |
|---|---|---|---|---|---|
| 2. Symphysis length 54.1 | 42.7 | 49.4 | 56.1 | 62.8 | 57.7 |
| 3. Maximal width of symphysis 29.8 | 27.7 | 34.6 | 42.2 | 39.8 | 29.3 |
| 4. Minimal width of symphysis 24.4 | 23.9 | 27.9 | 31.3 | 29 | 22.2 |
| 5. Maximal width between two rami 65.1 | 64.4 | ca.68 | ca.61 | ||
| 6. Height behind p2 27.7 | 24.5 | 26.9 | 29.2 | 32 | 29.4 |
| 7. Height behind m1 26.0 | 23.8 | 30.6 | 27.6 | 30.5 | 27.1 |
| 8. Height behind m3 | 33.7 | 34.7 | 40.3 | 32.6 | |
| 9. Height at coronoid process ca.70.0 | >56.6 | 79.5 | 80.3 | ||
| 10. Height at condyle 72.6 | 57.6 | 84.3 | 75.7 | ||
| 11. p1-p4 length 42.5 | 44.9 | ca.44 | 44.8 | 43.7 |
TABLE 3. Measurements of the upper incisors and canines (in mm) of Chleuastochoerus from the Linxia Basin.
| I1 | I2 | I3 | C | ||||
|---|---|---|---|---|---|---|---|
| L×W | L/W | L×W | L/W | L×W | L/W | L×W | L/W |
| C. linxiaensis sp. nov. | |||||||
| V18063 View Materials l | 23.0×11.6 | 1.98 | |||||
| r | 21.8×11.6 | 1.88 | |||||
| V 18064.1 l 8.0×7.0 | 1.14 | 7.5×4.6 | 1.63 | 15.2×7.8 | 1.95 | ||
| r 7.7×7.4 | 1.04 | 15.0×8.3 | 1.81 | ||||
| HMV 0 575 l | *4.5×2.8 | *1.61 | 6.3×3.6 | 1.75 | |||
| r *5.4×4.5 | *1.20 | 6.0×3.8 | 1.58 | ||||
| C. stehlini | |||||||
| V 18061 View Materials l 7.7×7.8 | 0.99 | 7.4×4.6 | 1.61 | 6.9×3.6 | 1.92 | 26.2× | |
| r 7.9×7.8 | 1.01 | 7.2×4.8 | 1.5 | 6.4×3.9 | 1.64 | 23.6× | |
| HMV 0 569 l 7.2×9.4 | 0.77 | ||||||
| r 6.6×5.9 | 1.12 | 6.0×3.9 | 1.54 | ||||
| HMV 1439 l 6.5×7.1 | 0.92 | 6.1×4.6 | 1.33 | 6.9×4.1 | 1.68 | 19×11.9 | |
| r 6.6×6.5 | 1.02 | 6.4×4.9 | 1.31 | 6.9×4.2 | 1.64 | 17×11.4 | |
| HMV 0 156 l | *4.5×2.6 | *1.73 | 7.2×4.1 | 1.76 | |||
| r | *5.0×2.7 | *1.85 | 7.6×4.2 | 1.81 |
TABLE 4. Measurements of the lower incisors and canines (in mm) of Chleuastochoerus from the Linxia Basin.
| i1 | i2 | i3 | c | |
|---|---|---|---|---|
| L×W | L/W L×W | L/W L×W | L/W | Li×La×Po |
| C. linxiaensis sp. nov. | ||||
| V 18064.2 l 4.0×5.0 | 0.80 5.2×6.0 | 0.87 4.3×4.3 | 1.00 | 8.3×5.5×5.6 |
| r 3.9×5.3 | 0.74 | 8.4×5.9×6.2 | ||
| HMV 0 575 l *3.4×3.2 | *1.06 *4.0×4.5 | *0.90 | ||
| r *3.5× | *3.2×4.3 | |||
| C. stehlini | ||||
| V 18065 View Materials l 3.9×5.8 | 0.67 5.8×6.5 | 0.89 | 11.6×6.2×9.6 | |
| r 3.8×5.8 | 0.66 5.4×6.1 | 0.89 5.0×4.6 | 1.09 | 9.1×6.8×8.3 |
| V18066 View Materials l 4.0×6.1 | 0.66 5.6×6.6 | 0.85 | 13.2×7.9×10.6 | |
| r 3.7×5.8 | 0.64 | |||
| V18067 View Materials l 3.8×5.6 | 0.68 6.0×7.0 | 0.86 | 12.9×6.4×11.6 | |
| r 4.2×5.8 | 0.72 6.1×7.0 | 0.87 | 13.2×7.2×11.6 | |
| V 18068 View Materials l 3.9×5.2 | 0.75 5.8×6.0 | 0.97 5.3×4.3 | 1.23 | 11.8×7.1×9.7 |
| r 3.9×5.0 | 0.78 5.8×6.0 | 0.97 4.9×4.6 | 1.07 | 12.2×7.2×9.7 |
| HMV 0 564 l 3.9×5.7 | 0.68 6.0×6.3 | 0.95 5.3×4.2 | 1.26 | 7.8×6.3×6.1 |
| r 3.9×5.6 | 0.70 5.9×6.5 | 0.91 5.2×4.3 | 1.21 | 8.0×6.0×6.3 |
| HMV 0 565 l 3.4×5.8 | 0.59 4.9×7.1 | 0.69 | 12.5×6.3×10.3 | |
| r 3.3×5.2 | 0.63 4.7×6.7 | 0.70 5.7×3.6 | 1.58 | 11.8×6.3×10.1 |
| HMV 0 566 l 4.0×5.8 | 0.69 5.7×7.0 | 0.81 5.9×4.8 | 1.23 | 13.0×6.6×11.4 |
| r 3.7×5.7 | 0.65 | 12.0× | ||
| HMV 1439 l 3.8×5.0 | 0.76 5.4×5.8 | 0.93 5.3×4.6 | 1.15 | 10.1×5.2×8.3 |
| r 3.9×5.3 | 0.74 5.9×6.2 | 0.95 5.0×4.6 | 1.09 | 9.8×5.0×8.4 |
| IVPP |
Institute of Vertebrate Paleontology and Paleoanthropology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.