
Ceratophrys aurita Raddi
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5094.3.9 |
|
publication LSID |
lsid:zoobank.org:pub:0233E081-5C5C-486B-8A20-4ED7B6BAB053 |
|
DOI |
https://doi.org/10.5281/zenodo.6302982 |
|
persistent identifier |
https://treatment.plazi.org/id/677187DE-924E-FFB4-6CBC-4F6F0D69FC90 |
|
treatment provided by |
Plazi |
|
scientific name |
Ceratophrys aurita Raddi |
| status |
|
Ceratophrys aurita Raddi View in CoL
Referred Specimen. ZUFABC 038, partial skull.
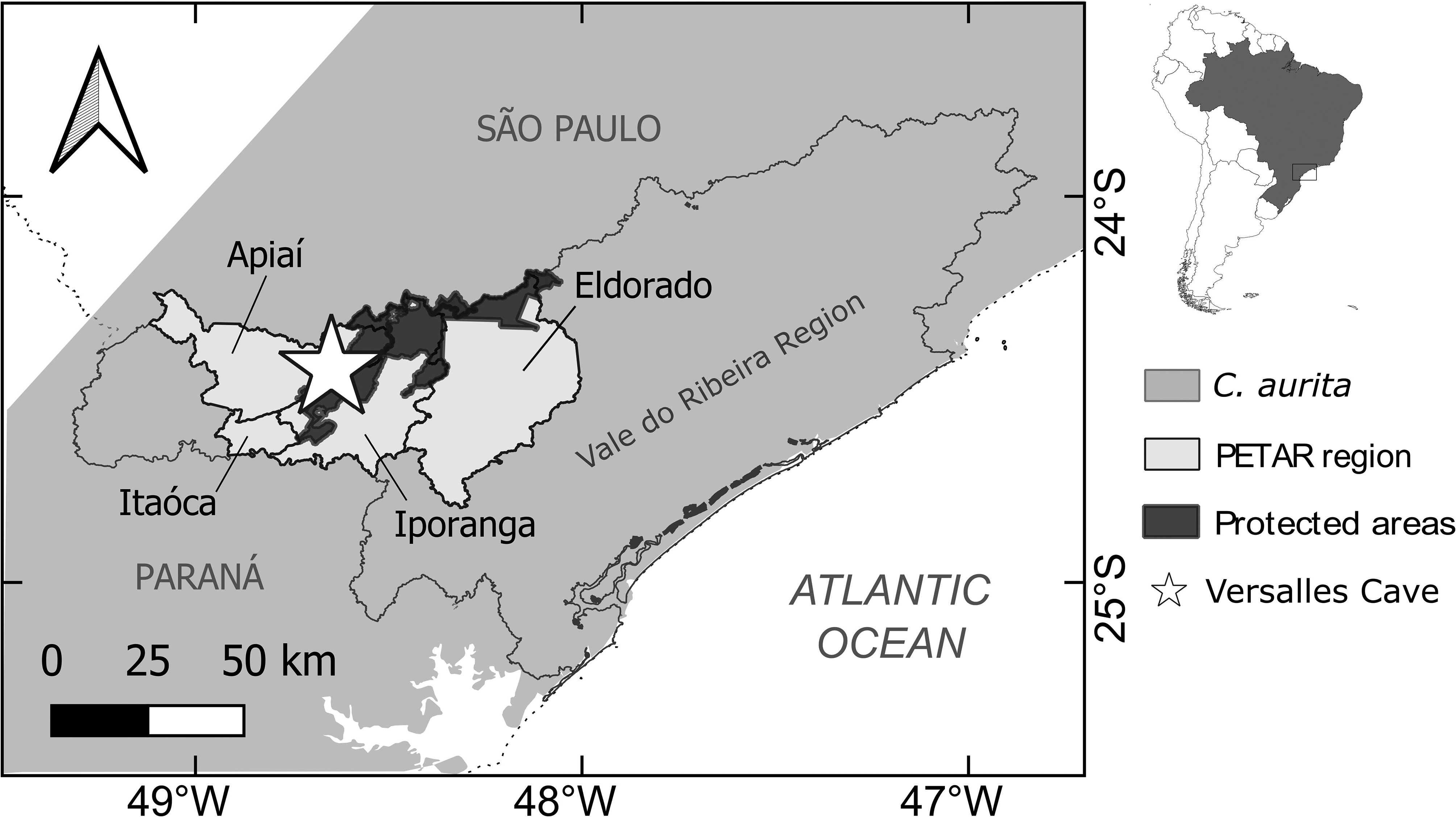
Locality and Age. Versalles Cave, Apiaí municipality, state of São Paulo, Brazil, Late Pleistocene–Early Holocene.
Distribution. The extant species distribution ranges from states of Minas Gerais and Bahia to Rio Grande do Sul, Brazil ( Frost 2021). The paleodistribution is restricted to one locality in the state of Minas Gerais ( Günther 1859 [ Barcelos et al. 2020]) and one locality in the state of São Paulo, Brazil.
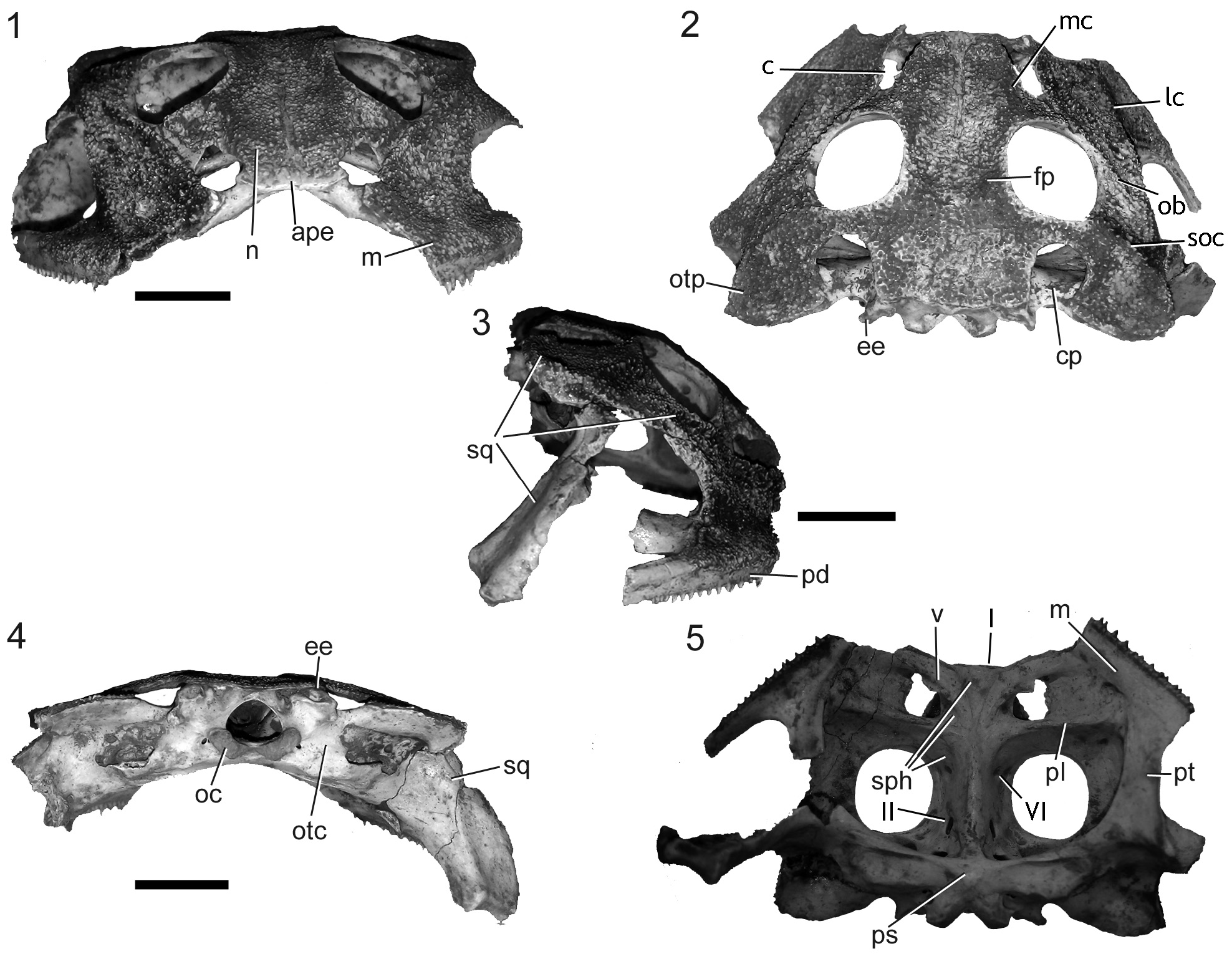
Description. The specimen ZUFABC 038 consists of a skull partially preserved and five fragments that were glued together. The left ventral ramus of squamosal, posteriormost and anteriormost portions of maxilla, premaxillae, left quadratojugal, septomaxillae and columellae are not preserved. The larger fragment consisted of the main parts of the skull. The hyperossification of the skull, with synostosis of skull bones are indicative of an adult specimen. The right maxilla is broken in three fragments from the skull, a cranial portion of the right pars facialis, including a fragment of the palatine and a fragment of the maxilla from region of the pterygoid process, with non-pedicellated fang-like teeth preserved. The right ventral ramus of squamosal synostosed to the quadratojugal constitute other two fragments. Those fragments were glued together ( Fig. 2 View FIGURE 2 ), and allowed the estimation of the approximate maximum cranium height (25 mm). We measured the width of the skull-half (from right quadrate to the jugular foramen = 30 mm) and mirrored it to infer the approximate maximum width (60 mm).
The nasals are paired bones that are flat, symmetrical and wide ( Fig. 2.1 View FIGURE 2 ). The nasals articulate synostotically with each other medially and bear exostosis in its dorsal surface, except throughout the axis of the suture between the nasals. The minimum-interorbital width is 9 mm, the maximum bi-orbital width is 29 mm, and the diameter of anteroposterior orbit is 10 mm. The nasals articulate synostotically with each frontoparietal posteriorly. The maxillary process of the nasals is bar-shaped and bear a medial crest on its proximal portion ( Fig. 2.2 View FIGURE 2 ). It articulates with each zygomatic ramus of the squamosal laterally, forming the infraorbital flange. The nasals, maxilla, and zygomatic ramus of the squamosal participates in the formation of the choanae aperture, which is oblong and directed medially. Frontoparietals are paired, symmetrical, dorsally exostosed bones, synostotically fused to each other throughout their contact. Each frontoparietal sutures laterally with the squamosal through a parieto-squamosal arch, forming a postorbital fenestra (diameter = 4 mm) ( Nicoli 2019).
Squamosals are paired, triradiate bones that bear exostosis on its dorsal surface. The three rami of the squamosal are: the ventral ramus, otic ramus, and zygomatic ramus. The squamosal otic ramus is exostosed dorsally and bear a subquadrangular otic plate. This otic plate is dorsolaterally curved and presents a dorsal concavity. The otic plate extends posteriorly beyond the level of the occipital condyles. The lateral portion of the otic plate also bears a lateral process ventrally projected. The maximum width between otic plates is 44 mm. The squamosal zygomatic ramus articulates with the pars facialis of the maxilla, with the maxillary process of the nasal and with the anterior ramus of the pterygoid. The ventral and zygomatic rami articulate with the maxilla and contribute to the formation of the subtemporal fenestra ( Fig. 2.3 View FIGURE 2 ). Two crests originate from the contact of the zygomatic ramus with adjacent bones: the lateral crest (that extends throughout the subtemporal fenestrae edge) and the oblique crest (that extends through the anterior-most edge of the orbit to the lateral process of otic plate. The squamosal ventral ramus constituted two fragments. It is an elongate element, ventrally directed, which sutures the quadrate and the pterygoid ( Fig. 2.4 View FIGURE 2 ).
Pterygoids are paired, triradiate bones. The anterior and posterior rami of the right pterygoid is heavily fragmented. The anterior and medial rami of left pterygoid are well preserved. The pterygoid anterior ramus sutures to the respective maxilla and palatine forming the lateral edge of the interpterygoid vacuity. The maximum inter-pterygoids distance is 22 mm. The pterygoid medial ramus sutures the parasphenoid and the otoccipital ( Fig. 2.5 View FIGURE 2 ). The posterior ramus sutures the ventral ramus of the squamosal and the quadrate. Only the right quadrate is preserved. The maxilla is preserved only between the vomer and the interpterygoid vacuity. The pars facialis of the right maxilla was split into three fragments. The pars dentalis of the maxilla bear fang-like non-pedicellate teeth (15 on the right and 6 on the left maxilla).
The sphenethmoid sutures the cultriform process of the parasphenoid posteriorly, the vomer and the palatine medially, and the nasals anteriorly. The anterior portion of sphenethmoid is a wide and truncated projection that bears two olfactory nerve foramina (CN I). A well-developed abducens nerve foramen (CN VI) is placed at the medial portion of sphenethmoid, on the suture with the cultriform process of parasphenoid. At the posterior portion, the sphenethmoid meets the otoccipital and the cultriform process of parasphenoid and forms the optic nerve foramen (CN II). The vomers are paired bones sutured to the sphenethmoid, and the maxilla. Vomers present vomerine teeth. The palatines are elongated paired bones. The parasphenoid is a T-shaped bone. The cultriform process of the parasphenoid does not reach the level of the planum antorbitale. The otoccipital sutures synostotically to the parasphenoid, the sphenethmoid, the frontoparietal, the squamosal, and the pterygoid. The otoccipital bears epiotic eminences flat in posterior view and slender in dorsal view. The epiotic eminences are partially overlapped by the processus posterior of the frontoparietal, forming the postemporal fenestra. The posterior portion of otoccipital bears juxtaposed occipital condyles.
The specimen ZUFABC038 is attributed to Ceratophryidae on the basis of the following synapomorphies: exostosed skull, parieto-squamosal arch present, zygomatic ramus of squamosal in contact with the maxilla, non-pedicellate and fang-like teeth, parieto-squamosal arch present, and the otic ramus of squamosal expanded and overlapping the prootics ( Lynch 1971, 1982; Fabrezi 2006; Fabrezi & Quinzio 2008; Nicoli 2019). ZUFABC038 is assigned to Ceratophrys by nasal with robust bar-shaped maxillary processes, parieto-squamosal arch forming a postorbital fenestra, absence of pars palatina (and palatine process) on the anterior half of the maxilla ( Nicoli 2019), the presence of posterior margin of dermocranium indented, and synostosis of most skull bones into an akinetic unit ( Wild 1997; Lynch 1982).
The specimen ZUFABC038 can be assigned to the [ C. aurita – C. joazeirensis ] clade by the presence of a medial crest on the nasal, presence of vomerine teeth, and lateral crests on the zygomatic ramus of squamosal and maxilla ( Nicoli 2019). Unfortunately, other characters diagnosing the [ C. aurita – C. joazeirensis ] and the southeastern clade (i.e. C. aurita species group) as a bony dorsal shield ( Nicoli 2019) are not preserved in the specimen ZUFABC038. Our comparative study on specimens character variation within Ceratophrys (Appendix 1) show that it is possible to diagnose the specimen ZUFABC038 as a fossil representative of C. aurita by sharing with other exemplars of this species the combination of characters: zygomatic ramus of the squamosal wide (narrow-shaped in C. joazeirensis ; Barcelos et al. 2020); three crests and two depressions on zygomatic ramus of squamosal and maxilla ( Fig. 2.2 View FIGURE 2 ) (absent in C. cranwelli and C. ornata ), the otic plate of the squamosal dorsally concave (convex in C. joazeirensis , C. cranwelli and C. ornata ); anterior-most portion of sphenethmoid forming a wide and truncated projection ( Fig. 2.1 View FIGURE 2 ) (short and rounded in C. joazeirensis ), and nasal and maxilla forming choanae aperture oblong-shaped ( Fig. 2.1 View FIGURE 2 ; Barcelos et al. 2020) (rounded in C. joazeirensis , C. cranwelli , and C. ornata ).
We are aware Nicoli (2019) hypothesizes C. aurita and C. joazeirensis may represent variations within a same species based on osteology. The author compared one specimen of C. aurita (MLP 1280) to two cleared and stained specimens of C. joazeirensis (MZUSP 56463, 126203) and claimed ontogenetic variation. Our sample includes eight C. aurita , and one specimen of C. joazeirensis , but despite observing some intraspecific variation in C. aurita , it is still possible to diagnose specimens as indicated above. Following our observations, we interpret the figures in Nicoli (2019) as follows: the morphology of the large specimen (MZUSP 56463) agrees with that diagnostic for C. aurita (i.e. three crests and two depressions on zygomatic ramus of squamosal and maxilla, anterior-most portion of sphenethmoid forming a wide and truncated projection, and nasal and maxilla forming choanae aperture oblong-shaped). The smaller specimen (MZUSP 126203) presented a rounded otic plate and zygomatic ramus of squamosal and maxilla without crests or depressions in its posterior edges, characters not congruent to [ C. aurita - C. joazeirensis ] clade, but it could be ontogenetic variation if it is a juvenile as Nicoli (2019) proposed.
Misidentification concerning C. aurita and C. joazeirensis are reported in literature ( Maciel et al. 2013), so it would be important to check for Nicoli’s specimens identities. Unfortunately, the information on locality or identity assertion of these exemplars are missing in Nicoli (2019). These specimens were not at MZUSP during our visit, and when we asked the collection manager for the data associated to MZUSP 56463 and MZUSP 126203, the answer was that the record of MZUSP 56463 was not available in the system, and that MZUSP 126203 was associated to a hylid originally from Werner Bokermann collection (WCAB). As the specimens studied by Nicoli (2019) are clearly Ceratophrys , it is something worth to check in the future.
We noted three polymorphic characters in the skull of C. aurita species based in our sample, but that not overlap with the characters observed in C. joazeirensis (under brackets): the ornamentation of the projection of the anterior portion of sphenethmoid can be wide and truncated (e.g. MZUSP 31367), or wide and rounded (e.g. KU 223818) (narrow and rounded); a concavity/scar on the suture between the vomer and the anterior portion of sphenethmoid may be present (e.g. cas:herp:84998) or absent (e.g. MZUSP 61114-2) (absent); Ceratophrys aurita ZUFABC 038 enlarges the range of skull–ratio for the clade [ C. aurita – C. joazeirensis ] from 41% (in C. aurita ZUFABC 038) to 50% (former skull-ratio proposed by Nicoli 2019) (50% in C. joazeirensis ). The intraspecific variation in the osteology of Ceratophrys species is unexplored, as for most anurans, and may highlight the discussion on paleodiversity and paleoclimatic changes in South America (e.g. Simon et al. 2016).
In contrast to the much better approached paleofauna of Ceratophryidae in Argentina, there is a hidden paleodiversity of Ceratophryidae in Brazil, with only four specimens registered (including the one in this paper). Ceratophrys aurita ZUFABC 038 is the third record of C. aurita and fourth record of Ceratophryidae for the Brazilian territory. The fossiliferous locality of Versalles Cave is located in the current distribution range of C. aurita ( Fig. 1 View FIGURE 1 ; Araújo et al. 2010). Still, the distribution of C. aurita during the last glacial maximum was potentially broader than the current one ( Vieira et al. 2018), considering the expansion of southeastern forests to the interior of the country. The record of the specimens Ceratophrys aurita BMNH 18895/6 from Lagoa Santa–MG is congruent to this more continental paleodistribution. Lastly, Ceratophrys aurita ZUFABC 038 states the extant species occurs in the southeastern of Brazil for at least the Late Pleistocene.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.