
Bombus trifasciatus, LINEAGE
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00861.x |
|
persistent identifier |
https://treatment.plazi.org/id/8F061705-FFB8-FF8D-FC1A-F9C8FDF4FA12 |
|
treatment provided by |
Marcus |
|
scientific name |
Bombus trifasciatus |
| status |
|
B. TRIFASCIATUS LINEAGE View in CoL
Several of the nominal taxa in the B. trifasciatus lineage that were described as species by early authors are resolved by DNA evidence with relatively old unique histories, including B. t. albopleuralis , B. t. malaisei , B. trifasciatus s.s., B. t. montivagus , and to a lesser extent B. t. wilemani and B. t. maxwelli . In addition our study identified a new most distant taxon within this lineage from western Myanmar (photo in Fig. S1 View Figure 1 ). This western Myanmar taxon will remain unnamed until males can be found to compare to the types of the nominal taxon B. t. magrettianus, which was described by Richards (1931) from Sikkim, perhaps in error ( Williams, 1991).
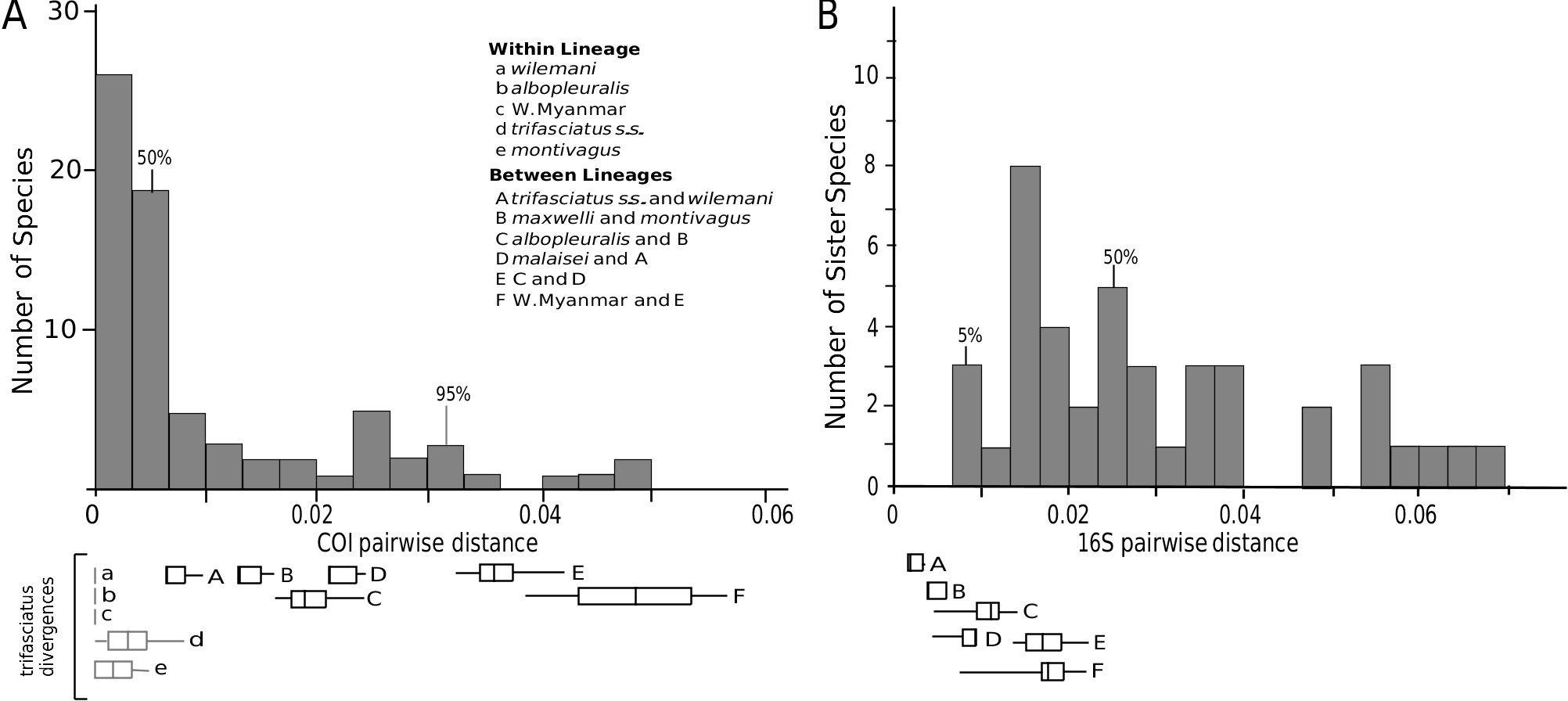
Several lines of evidence can be argued to support that the B. trifasciatus lineage might include several species. Several previously recognized nominal taxa in this lineage exhibit mitochondrial distances between them and sister sublineages that are within the range (and near the mode) of genetic distances typically found between bumble bee sister species. These lineages also have divergences that fall in the far right tail of those typical within bumble bee species, and in the case of the western Myanmar sublineage, even exceeding known intraspecific divergences. In contrast, within each of these sublineages, genetic distances are very low, representative of population-level divergences ( Fig. 7 View Figure 7 ). Nuclear ITS has fixed conflict-free haplotypes supporting several of the major sublineages, but does not distinguish amongst the nominal taxa B. t. albopleuralis , B. t. maxwelli , and B. t. montivagus , or distinguish B. t. malaisei from B. trifasciatus s.s. The nuclear genes ArgK and PEPCK and wing morphometrics show some clustering, but without more sampling it is hard to discern whether shared haplotypes between sublineages in these genes result from introgression or from maintained ancestral polymorphisms.
Previous arguments that these sublineages appear to be undergoing gene flow, and therefore represent a single species, are not supported by the genetic data. Williams (1991) synonymized the nominal taxa using the argument that several features of colour pattern appear to be intermediate where those taxa occur in proximity. For example, the black at the posterior of tergum 3 in the east Himalayan colour patterns is intermediate between the west Himalayan colour patterns and the more eastern colour patterns. However, our genetic results support all of the Himalayan colour patterns as part of a single sublineage, with no mitochondrial variation detected amongst sampled sequences. The B. trifasciatus s.s. and B. t. montivagus colour patterns represent separate sublineages with relatively large genetic distances from the Himalayan sublineage. It is possible, however, that the single colour pattern resembling the nominal taxon B. albopleuralis gantokiensis resolved within the B. t. montivagus sublineage could be an example of mitochondrial introgression between the B. t. montivagus and B. t. albopleuralis sublineages. Similarly, specimens resembling the B. trifasciatus s.s. colour pattern but with much more extensive black on the mesosoma (the nominal taxon Bombus minshanicus ) were allied to those with typical B. trifasciatus s.s. colour pattern in mitochondrial and nuclear genes, with no indication of introgression with the black mesosomal forms of the B. t. albopleuralis sublineage in our samples. Williams (1991) suggested that some colour patterns collected in Sichuan could be intermediate between other colour forms. One candidate for this would be the nominal taxon B. t. malaisei . Results from mitochondrial genes place this in its own distinct sublineage. Similarly, Williams (1991) noted that wing darkness lightens from west to east in the B. trifasciatus lineages. The results here show that, although this is true at the broad scale, wing colour tends to be lineage specific with no obvious signs of introgression.
Some of the reasons for intermediate colour traits where lineages converge geographically could be attributed to the mimetic process itself. Neighbouring mimicry groups may have more similar patterns than more geographically distant ones, because changing species assemblages between neighbouring areas across time will affect the establishment of local mimetic colour convergences. This is particularly likely when considering that the sampled colour patterns of comimics in the B. breviceps and B. haemorrhoidalis lineages are each more likely to be conspecific, as proposed by Williams (1991). Within these two species, gene flow could in principle occur between regions, potentially leading to intermediate colour forms that could be mimicked by the B. trifasciatus lineage through selection, even though the B. trifasciatus lineage may have little to no gene flow between its sublineages. Lastly, such colour patterns could result from long-maintained patterns from a time when gene flow between colour patterns in these regions did occur. Irrespective of the mechanism, finding intermediate mimetic colour patterns across a wide range may not necessarily be a good indication of gene flow between lineages in the present day.
Given the possibility that the B. trifasciatus lineage consists of several species, where might the species boundaries lie? This involves taking into consideration evidence for gene flow, including degree of lineage monophyly across genetic markers, genetic relative to geographical distances, and similarity in morphometrics and morphology. A more quantitative measure, however, is genetic divergence amongst mitochondrial markers. Genetic divergence as a criterion for species delimitation must be treated with caution, as this is only indirectly related to species concepts and levels of interspecific and intraspecific variation typically show substantial overlap (e.g. Meier et al., 2006). As a more conservative approach using this measure, we have compared the divergences in our results with the maximum intraspecific divergences observed amongst other bumble bees. These data, in combination with the lineage clustering observed using nuclear markers and wing morphometrics, support the following sublineages within the B. trifasciatus lineage as the most likely candidates for recognition as separate species: B. montivagus (including as subspecies: B. m. maxwelli and B. m. albopleuralis ), B. trifasciatus (including as subspecies: B. t. wilemani and B. t. malaisei ), and an as-yet unnamed taxon from western Myanmar. Comparing our results with typical interspecific divergences in Bombus provides a less conservative boundary on the number of species, suggesting that B. t. albopleuralis and B. t. malaisei might also be separate species. Additional support for these lineages includes low genetic variation in mitochondrial markers for B. t. albopleuralis relative to other sublineages and a lack of correlation between genetic distances and geographical distances involving these taxa. Sampling of additional nuclear markers and a more expansive population-level sampling regime in areas of sympatry, such as Yunnan, Sichuan, southern China, and northern Myanmar, will provide more definitive conclusions on species recognition.
Morphological features examined in females show only subtle diagnostic traits for the species recognized conservatively here, contributing to difficulties in making species decisions ( Williams, 1991; Williams et al., 2009). The first feature unrelated to colour used previously was relative malar space height to width ( Richards, 1931). Quantification of this trait across numerous specimens revealed it to be a poor indicator of sublineage affinity. The nominal taxon B. trifasciatus s.s., for example, exhibited variation in the relative length of the malar space across the full range of measurements in the B. trifasciatus lineage , with specimens in central China having the longest and those in eastern China having the shortest malar spaces. Corolla-tube lengths of the regional flora and the competition for these flowers by local pollinator communities might create different adaptive landscapes for tongue length (cf. Heinrich, 1979). Therefore, the length of malar space, which is positively correlated with tongue length, could be under strong local selection.
Owing to the current lack of strong diagnostic traits in examined females of the B. trifasciatus lineage , nominal taxa corresponding to sublineages must be distinguished for females by using colour pattern and geographical distribution. Males usually are often much easier to diagnose than females in bumble bees because of their rapidly evolving and morphologically complex genitalia ( Williams, 1991; Williams et al., 2009). Additional collection of male specimens from the sublineages may show traits for better morphological species diagnosis in the future.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |