
Xanthorhoini Pierce, 1914
|
publication ID |
https://doi.org/10.5281/zenodo.279481 |
|
DOI |
https://doi.org/10.5281/zenodo.6184106 |
|
persistent identifier |
https://treatment.plazi.org/id/D2566362-FF86-FFD1-FF75-FAD2DF5DF817 |
|
treatment provided by |
Plazi |
|
scientific name |
Xanthorhoini Pierce, 1914 |
| status |
|
Tribe Xanthorhoini Pierce, 1914
(Fig s 130–148)
The original treatment in Pierce (1914) included Xanthorhoe Hübner , Orthonama Hübner , Ochyria Hübner , Camptogramma Stephens, Percnoptilota Hulst and Mesotype Hübner but also Anticlea Stephens and Earophila Gumppenberg (both genera without coremata) and may be interpreted different as compared with preceding Epirrhoinae: Cöremata at seventh segment of male abdomen; valve costa greatly extended with projecting process; valvula reduced; calcar long with geniculate head (vestigial in some New Zealand taxa: Craw 1987); manica heavily sined; signum a pyriform band, consisting of overlapping spines; small-sized, heavily sclerotized male armatures.
Forbes (1948) stressed the characters of Xanthorhoini as follows: the straight, “erect” hind wing upper discocellular vein (mdcv, here DC1) and large coremata pencils on seventh abdominal segment of male (subsuming Euphyia Hübner into his genus Xanthorhoe Hübner ).
Herbulot (1961) recombined the groups Epirrhoinae Pierce and Ortholithinae Pierce in Xanthorhoini . Holloway (1997: 190) wrote: “The presence of a large pair of coremata just distal to the eighth segment in male abdomen appears to be the most constant feature for this tribe. Though the occurrence of a calcar ... is an additional diagnostic feature…”. Holloway (loc. cit.) diagnosed the calcar as a curved process, club-like structure arising centrally from the juxta between the bases of valves. The group requires a global revision ( Holloway et al. 1987).
Modifications in the region of fultura inferior as defined here, include: Sternite A7 is membranized to a filiform stripe, the corresponding tergite to a bow-shaped sclerite.
Sternite A8 is reduced to a stripe, smooth in Holarctic species but V-shaped angulate in many southern hemisphere genera ( Figs 136, 137 View FIGURES 136 – 138 ).
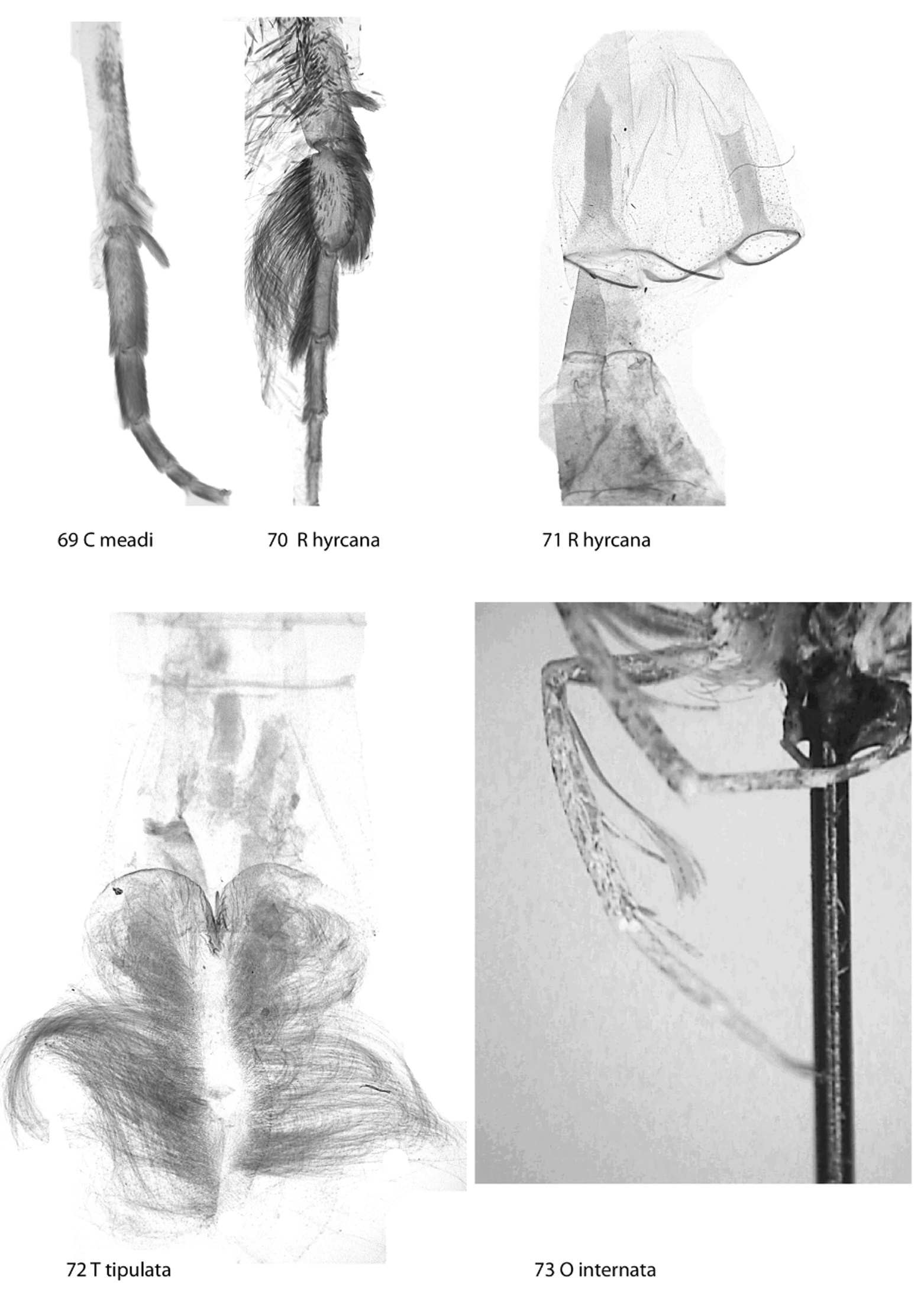
Tergite A8 usually consists of a basal stripe and dorsal semi-membranous flap ( Figs 131 View FIGURES 130 – 135 , 136 View FIGURES 136 – 138 ). In Rheumapterini , coremata are at the eighth segment, with both sternite and tergite dissclerotized to a slender central band ( Fig. 71 View FIGURES 69 – 73 ); labides sometimes present, on fultura, ( Craw 1987), or headed on stalk.
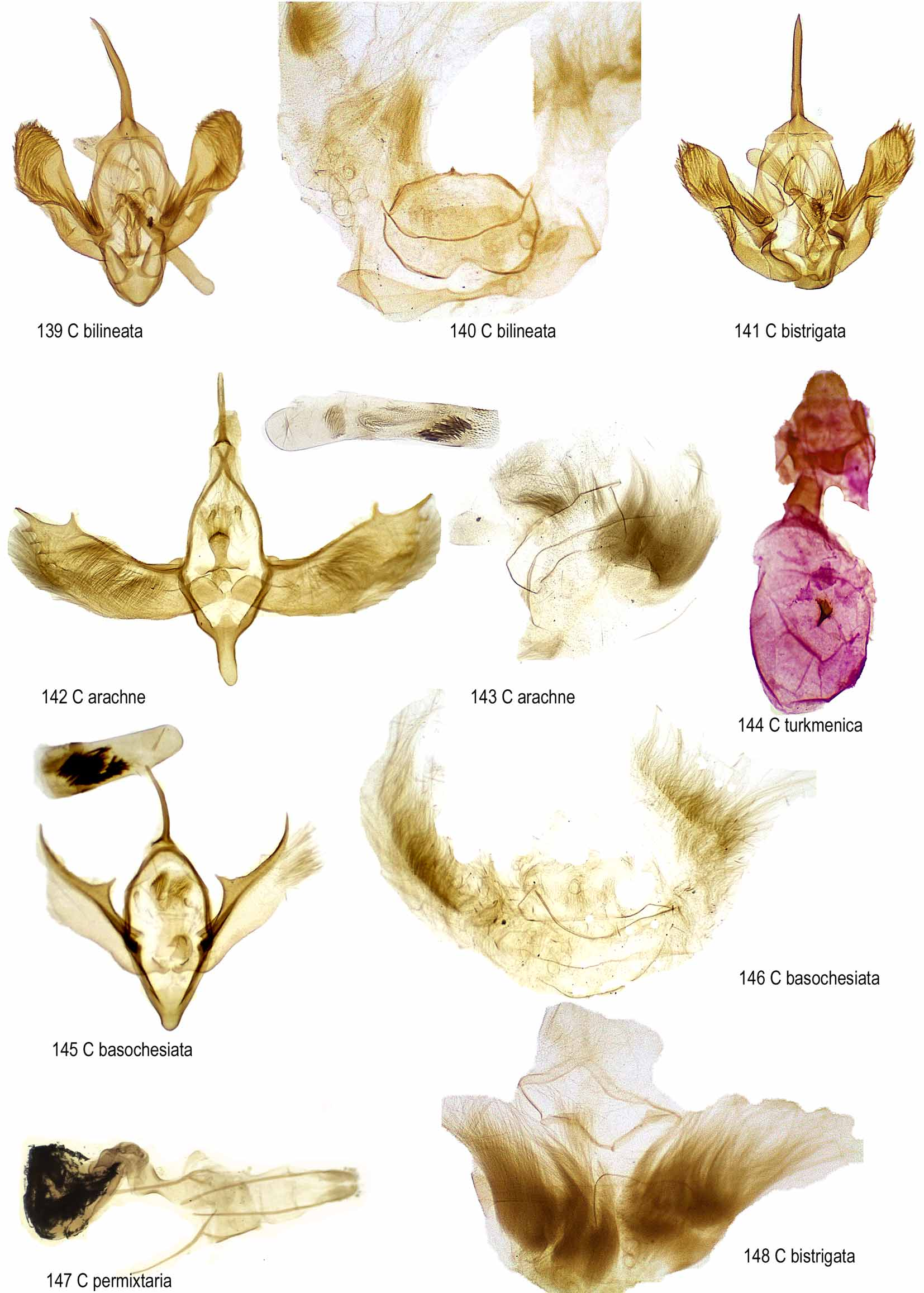
The tribe as defined above is polymorphic. The seventh segment may be broad, variously modifed both in south hemisphere and boreal genera ( Camptogramma Stephens Figs 139, 140 View FIGURES 139 – 148 ). Male genital armatures are large in some genera ( Catarhoe Prout , Figs 142–148 View FIGURES 139 – 148 , Epirrhoe , Fig. 153 View FIGURES 153 – 154 ). There are sclerotized longitudinal folds in epirrhoine and catarhoine female genitalia. Epirrhoina Pierce, 1914 was synonymized with Xanthorhoini by Herbulot (1961).
The presence of extensile coremata to the distal end of male abdomen, associated with membranization of the two last segments, is shared by Xanthorhoini , Cataclysmini, Euphyiini. The presence of a medial appendage to juxta (calcar) is shared by most xanthorhoine genera and the Larentiini but not Euphyiini. Female apophyses are short and weak in Xanthorhoini and Cataclysmini, but not in Euphyiini.
Distribution of Xanthorhoini : all regions, tending boreal and mountainous.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |