
Plecia parisiensis, Gee & Nel & Menier & De Ploëg, 2001
|
publication ID |
https://doi.org/10.5281/zenodo.5373084 |
|
publication LSID |
lsid:zoobank.org:pub:CE823B45-1F6E-47B8-A5EA-1C70028594F6 |
|
persistent identifier |
https://treatment.plazi.org/id/F3F16353-4201-4C2A-A801-9C67986A8EEC |
|
taxon LSID |
lsid:zoobank.org:act:F3F16353-4201-4C2A-A801-9C67986A8EEC |
|
treatment provided by |
Marcus |
|
scientific name |
Plecia parisiensis |
| status |
sp. nov. |
Plecia parisiensis View in CoL n. sp. ( Figs 1-4 View FIG View FIG View FIG View FIG )
TYPE MATERIAL. — Holotype: male specimen (PA 74) (2/5). Paratypes: male specimen (PA 74) (3/5), female specimen (PA 74) (4/5) (these three specimens were fossilised in the same cylindrical piece of amber, which was 2 cm long and 0.5 cm in diameter, together with an Hymenoptera and a male Diptera , Psychodidae Bigot, 1854 ); male specimen (PA 2530) (1/2) (fossilised with an adult Ephemeroptera), collection Gaël De Ploëg and Indivision Langlois-Meurinne, deposit- ed in the Muséum national d’Histoire naturelle, Paris . The amber pieces are mounted on microscope slides in glass cells, and embedded in Canada balsam.
ETYMOLOGY. — parisiensis is made after Paris Basin.
TYPE LOCALITY. — Le Quesnoy, Chevrière, region of Creil, Oise department, Paris Basin , France.
TYPE HORIZON. — Lowermost Eocene, Ypresian (Sparnacian), level MP 7 of the mammal fauna of Dormaal. It has been demonstrated before that this amber is native, and very different from the Baltic amber in age, chemical composition and origin ( Feugueur 1963; De Ploëg et al. 1998; Nel et al. 1999).
DESCRIPTION
Holotype specimen PA 74 (2/5)
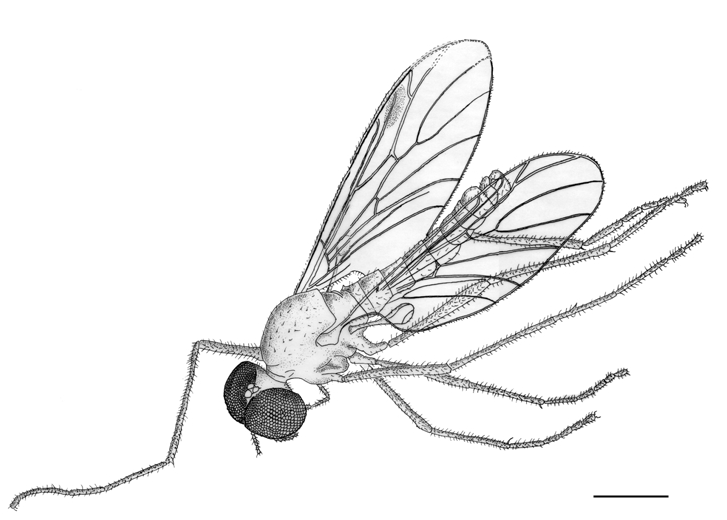
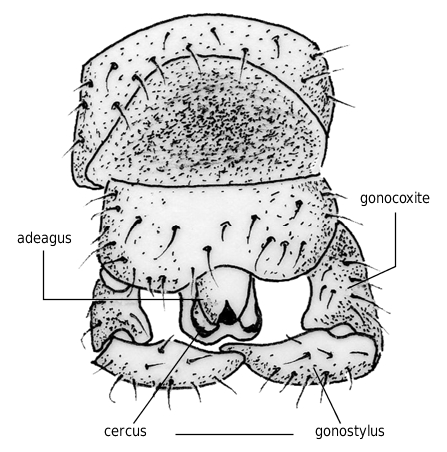
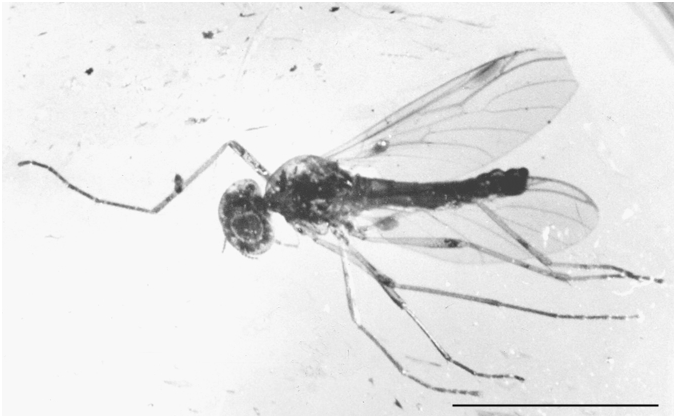
Body length 4.9 mm. Specimen complete ( Fig. 2 View FIG ), except for the apex of the right forewing which is lacking. Eyes large and dorsally merging (male bibionid character). The three ocelli (diameter 0.1 mm each) are on a dorsal protuberance. Antennae with 10 antennomeres. Thorax gibbous, with sparse short dorsal setae, 1.4 mm long and 0.9 mm wide. Fore legs 4.7 mm long, median legs 3.85 mm long, and hind legs 4.8 mm long. Legs show no strong spine. For the detailed dimensions of the legs, see Table 1. Abdomen narrowly elongated, 2.65 mm long and 0.5 mm wide. Wings ( Fig. 3 View FIG ): forewings rather narrow, 4.25 mm long and 1.45 mm wide (i.e. about 3.5 times longer than wide). Anal lobe well-developed. Vein R2+3 short, strongly oblique, making an angle of 30° with R4+5. R2+3, 0.38 mm long. R4+5, 0.96 mm long. Costal vein C ending at the apex of the wing, 0.3 mm distad of the apex of R4+5. Vein R1 reaching the wing margin halfway between Sc and R2+3. Pterostigma elliptic, brown, 0.58 mm long and 0.19 mm wide. Crossvein r-m perpendicular to M and Rs. The distance between r-m and the fork of M is one fourth the length of M1. M1, 1.56 mm long, M2, 1.27 mm long. Wing hyaline. The notation of the points used here under to describe the wing venation are after Melander (1949: 18, fig. 1). Distances A-B 0.66 mm; A-C 1.46 mm; A-M 0.25 mm; B-G 0.22 mm; B-J 0.19 mm; C-D 0.10 mm; C-E 0.20 mm; D-H 0.19 mm; M-J 0.54 mm; M-K 0.88 mm; O-N 0.58 mm. Halters 0.85 mm long, ovoid with a blunt apex, brown in color ( Figs 1 View FIG ; 2 View FIG ). Genital appendages ( Fig. 4 View FIG ): gonostyli long ( 0.23 mm) and straight, with an obtuse apex. Cerci weakly developed. Ninth tergum 0.4 mm wide. It bears a strong median indentation.
Paratype specimen PA 74 (3/5)
Body length 3.9 mm. Specimen incomplete. The head and the wings are partly destroyed. This specimen is distinctly smaller than the holotype but it has the same general habitus. Fore legs 3.8 mm long, median legs about 3.4 mm long, and hind legs about 4.0 mm long. As in males, the legs show no strong spine. Abdomen narrowly elongated, 2.4 mm long and 0.4 mm wide. Wings ( Fig. 3 View FIG ): forewings 3.8 mm long. Anal lobe well-developed. Vein R2+3 short, strongly oblique, making an angle of 30° with R4+5. R2+3, 0.20 mm long. Costal vein C ending at the apex of the wing, 0.2 distal of the apex of R4+5. Vein R1 reaching the wing margin halfway between Sc and R2+3. Pterostigma elliptic, brown, 0.5 mm long and 0.1 mm wide. Wing hyaline. Halters identical to those of the holotype. Genital appendages poorly preserved.
Female paratype specimen PA 74 (4/5)
Body length about 4.5 mm. Specimen incomplete. The head and the wings are partly destroyed. This specimen is almost of the same size as the holotype and it has the same general habitus, except for the broader abdomen. Forewings hyaline, 4.2 mm long. The wing venation is identical to that of the male holotype. Abdomen broader than that of the male, 3.3 mm long and 1.1 mm wide. The genital appendages are retracted in the abdomen, then poorly visible.
Male paratype specimen PA 2530 (1/2)
Body length about 3.9 mm. Specimen complete, but its genital appendages are hidden by the wings. This specimen has almost the same size and habitus as the paratype PA 74 (3/5). Forewings hyaline, 3.7 mm long.
DISCUSSION
This species is attributed to the genus Plecia Wiedemann, 1828 because of the simple legs, i.e.
without large apical spur or spine, veins Rs furcated and R2+3 oblique and short.
About 150 species of fossil Plecia have been described from lacustrine Cenozoic sediments, but most of them are rather badly known because of poor descriptions and/or figures. A revision of these fossil species would be necessary. In most descriptions the genital appendages and body structures are not figured and not properly described by the authors ( Oustalet 1870; Théobald 1937; Zhang et al. 1994 among others). The type specimens of some species were originally in a very poor state, especially the material described by Oustalet (1870), preserved in MNHN, Paris .
Because of the lack of phylogenetic analysis of the Pleciinae that includes the fossil species, we cannot determine the exact affinities of Plecia parisiensis n. sp. We have compared Plecia parisiensis n. sp. to the Plecia species described from the Upper Cretaceous or Eocene lacustrine deposits. These later descriptions are mainly based on the wing venation. Plecia parisiensis n. sp. differs mainly from these taxa because of its smaller dimensions, all the Eocene Plecia being larger with longer and wider wings.
More precisely, Plecia parisiensis n. sp. differs from Plecia undans Zeuner, 1941 (upper Paleocene/ Eocene, Ardtun Beds, Isle of Mull, Scotland, UK) in its distinctly smaller dimensions (wing lengths range 3.7-4.25 mm against more than 10.3 mm in
A new Plecia sp. ( Insecta, Diptera , Bibionidae ) in lower Eocene amber
P. undans View in CoL ). Plecia akerionana Fitzgerald, 1999 View in CoL (Eocene, Green River Formation, Wyoming, USA) has smaller wings ( 3.47 mm long), light brown fumose. Plecia rhodopterina Cockerell, 1924 View in CoL (Eocene, Green River Formation, Wyoming, USA) has wings of nearly the same length ( 5 mm) but they are dark ferruginous, R2+3 is distinctly longer. Plecia pealei Scudder, 1890 View in CoL and Plecia dejecta Scudder, 1890 View in CoL (Eocene, Green River Formation, Wyoming, USA) have longer wings (respectively 10 mm and 7.5 mm long). Plecia woodruffi Cockerell, 1916 View in CoL (Eocene, Green River Formation, Ute Station, Utah, USA) has longer wings ( 7 mm long) with the costal area suffusedly dusky. Plecia winchesteri Cockerell, 1917 View in CoL (Eocene, Colorado, USA) also has longer wing ( 7.3 mm long) and “dilute fuliginous” ( Scudder 1890; Cockerell 1916, 1917). Plecia minutula Rice, 1959 View in CoL (Eocene, British Columbia, Canada) has wing slightly longer ( 5.5 mm long) with the apex distinctly more rounded than P. parisiensis View in CoL n. sp. All the other species of Plecia View in CoL described by Scudder (1890), Handlirsch (1910) and Rice (1959) from the Eocene of British Columbia have distinctly longer wings (more than 5.5 mm long). Schröder (1999) and Rust (1999) described and figured a Plecia sp. and a Penthetria sp. from the Paleocene/Eocene of the Fur-Formation ( Denmark). Both have a vein R2+3 more or less straigth, very different of that of P. parisiensis View in CoL n. sp. Plecia acourti Cockerell, 1921 View in CoL (upper Eocene, Isle of Wight, UK) has longer wings ( 6.4 mm) entirely dark fuliginous ( Cockerell 1921).
All the Plecia View in CoL described by Théobald (1937) from the upper Eocene of the Gard department ( France) have longer wings, ranging between 6.0 mm to 10 mm long.
Plecia fushunensis View in CoL ( Hong 1980: 47; 48, textfig. 18, pl. 1; fig. 3) was described as Lacibibio fushunensis View in CoL , and renamed Plecia View in CoL by Zhang (1989). Unfortunately, the successive drawings of the wings are unexploitable. After Evenhuis (1994), this fossil is supposed to originate from the Chinese Eocene amber of Fushun, but it appears to be clearly an impression of an isolated wing on a lacustrine rock, after the original photograph of Hong. This wing clearly differs from Plecia parisiensis View in CoL n. sp. in its very long and oblique R2+3, intermediate in shape between those of the modern species of Plecia View in CoL and Penthetria View in CoL . It is also about 9.5 mm long.
Table 2 compares the few species described from different ambers. It clearly shows that our new species cannot be one of these already described taxa. In particular, Plecia parisiensis View in CoL n. sp. cannot be attributed to any of the species of Plecia View in CoL described from Baltic amber. The original descriptions and figures of Plecia prisca Meunier, 1899 View in CoL and Plecia borussica Meunier, 1907 View in CoL are very unclear. Nevertheless, Plecia borussica View in CoL differs from P. parisiensis View in CoL in having 10 antennomeres and its third antennomere drop-shaped, this making it different from all others ( Meunier 1899, 1907). This once again corroborates our previous observations that the entomofauna of Le Quesnoy shows great differences when compared to the Baltic amber.
MISCELLANEOUS REMARKS AND NEW SYNONYMIES Oustalet (1870) described Bibio edwardsii from the Oligocene of Corent (Puy-de-Dôme, France). Evenhuis (1994) attributed it to the genus Penthetria Meigen, 1803 . After a thorough reexamination of the type specimen preserved in the collection of the Laboratoire de Paléontologie, Muséum national d’Histoire naturelle, Paris (specimen MNHN-LP-R.06669), it is clearly visible that vein R2+3 is short and not parallel to R4+5. Thus, we reattribute this species to the genus Plecia , under the name Plecia edwardsii ( Oustalet, 1870) n. comb.
Oustalet (1872) described an adult bibionid from the Eocene of the “Gypse de Paris ” (Buttes- Chaumont, Paris , France), under the name Bibio chapuisii . This species is not listed by Evenhuis (1994). We could not find the type specimen in the collections of the Paris Museum. Nevertheless, it is clear after the original description of Oustalet who indicated that this fossil is nearly identical to Plecia edwardsii ( Oustalet, 1870) , that B. chapuisii belongs to the genus Plecia . We propose to rename it Plecia chapuisii ( Oustalet, 1872) n. comb. The wing of P. chapuisii is 6.25 mm long, distinctly longer than that of P. parisiensis n. sp.
Penthetria oustaleti (Brongniart, 1876) View in CoL , from the Oligocene of Chadrat (Puy-de-Dôme, France), described under the name Protomyia oustaleti by Brongniart (1876a, b), has been attributed to the genus Penthetria View in CoL by Evenhuis (1994) who apparently ignored the revision of Brongniart (1878) who reattributed it to the genus Plecia View in CoL . A thorough reexamination of the type specimen, preserved in the collections of the Laboratoire de Paléontologie, Muséum national d’Histoire naturelle, Paris View in CoL (specimen MNHN-LP-R.06698), showed that it is a true Plecia View in CoL . Thus, we restore it into this genus, under the name of Plecia oustaleti (Brongniart, 1876) stat. rest.
PALAEOECOLOGICAL CONSIDERATIONS
The insect fauna of Le Quesnoy shows a great number of Diptera , mainly Nematocera. On a total of 10 000 fossil insects identified today from this location, four Bibionidae specimens have been found, making this family relatively more frequent in this amber than in any other amber (only two specimens described from the very rich fauna of the Baltic amber). The genus Plecia is very frequent in the upper Eocene, Oligocene and Miocene lacustrine sediments of western Europe, but it seems somewhat less frequent in the Paleocene and lower Eocene lacustrine sediments of the same areas (Menat, Messel) where, nevertheless, they still remain more frequent than in ambers. The origin of this difference between ambers and lacustrine outcrops is still questioned. One hypothesis could be that the Bibionidae are usually larger than other flies and thus more difficult to preserved in resins. Tipulids are distinctly larger than bibionids but also more frequent in Baltic and Oise ambers. Thus, this hypothesis is clearly not sufficient. Another one could be that the Bibionidae are not attracted by resins, similarly to what Skalski (1976) demonstrated for the some Lepidoptera, but this would need confirmation after experimentation on living taxa. A last hypothesis is that “march flies” appear quite early in the season, perhaps when resin production is at a low level. This shift in periods of appearance could be a reason why bibionids are so rare in ambers.
Bibionidae View in CoL larvae feed on “decaying organic material” and “root of grasses and other plants” ( Hardy 1986), in wet environments, which fits with the reconstruction of the palaeoenvironment proposed by Nel et al. (1999).

| MP |
Mohonk Preserve, Inc. |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Plecia parisiensis
| Gee, Julien, Nel, André, Menier, Jean-Jacques & De Ploëg, Gaël 2001 |
P. parisiensis
| Gee & Nel & Menier & De Ploëg 2001 |
P. parisiensis
| Gee & Nel & Menier & De Ploëg 2001 |
Plecia parisiensis
| Gee & Nel & Menier & De Ploëg 2001 |
Plecia parisiensis
| Gee & Nel & Menier & De Ploëg 2001 |
P. parisiensis
| Gee & Nel & Menier & De Ploëg 2001 |
Plecia akerionana
| Fitzgerald 1999 |
Lacibibio fushunensis
| Hong 1980 |
Plecia minutula
| Rice 1959 |
P. undans
| Zeuner 1941 |
Plecia rhodopterina
| Cockerell 1924 |
Plecia acourti
| Cockerell 1921 |
Plecia winchesteri
| Cockerell 1917 |
Plecia woodruffi
| Cockerell 1916 |
Plecia borussica
| Meunier 1907 |
Plecia borussica
| Meunier 1907 |
Plecia prisca
| Meunier 1899 |
Plecia pealei
| Scudder 1890 |
Plecia dejecta
| Scudder 1890 |
Bibionidae
| Macquart 1838 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Plecia
| Wiedemann 1828 |
Penthetria
| Meigen 1803 |
Penthetria
| Meigen 1803 |