
Cambarus loughmani Foltz, Sadecky, Fetzner and Thoma, 2019
|
publication ID |
https://doi.org/ 10.1080/00222933.2018.1557271 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD1443-E412-FFA8-C926-FE2DB94FF9C1 |
|
treatment provided by |
Felipe |
|
scientific name |
Cambarus loughmani Foltz, Sadecky, Fetzner and Thoma |
| status |
sp. nov. |
Cambarus loughmani Foltz, Sadecky, Fetzner and Thoma View in CoL sp. nov.
Cambarus dubius Faxon 1884: 114 View in CoL [in part]. Faxon 1885: 70, pl. 4: fig. 3; pl. 8: figs 7, 7 ʹ [in part]; Dewees 1972: 1, figs 1b–l, 2c–h, 3, 4, 5b–h, 6–16 [in part].
Cambarus carolinus dubius View in CoL .– Faxon 1914: 396, 425 [in part].
Cambarus (Jugicambarus) dubius View in CoL .– Hobbs and Bouchard 1973: 62 [in part]; Hobbs 1974: 18, fig. 60 [in part]; Hobbs 1989: 22, fig. 78 [in part]; Jezerinac et al. 1995: 121, fig. 61 [in part].
Cambarus aff. dubius View in CoL . – Loughman et al. 2015: 534 [in part].
Diagnosis
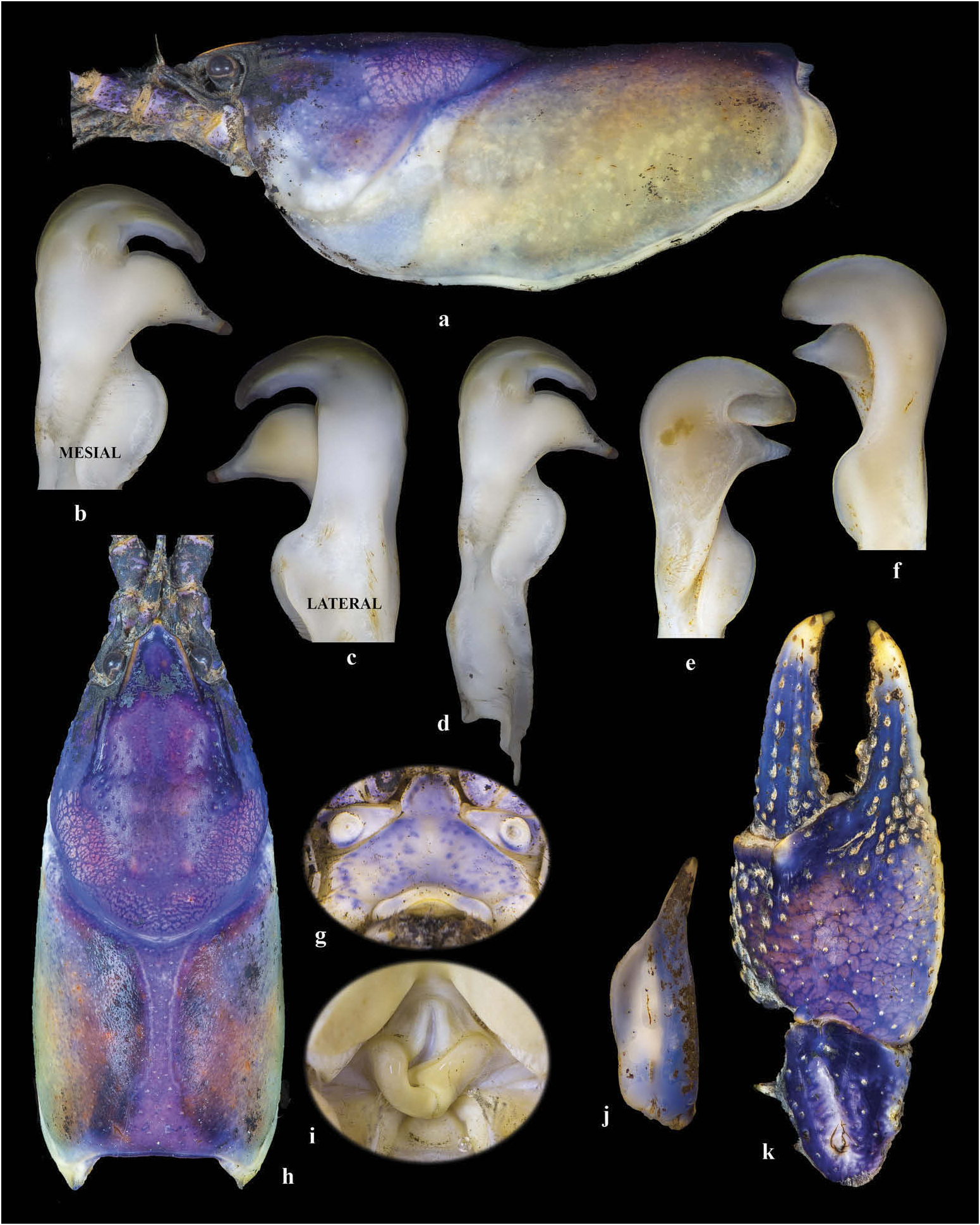
Body and eyes pigmented ( Figure 3 View Figure 3 ). Posterior region of rostrum concave, deflected anteriorly. Rostral margins thickened, running near parallel before converging at acumen. Rostrum moderately excavated. Acumen triangular terminating in distinct upturned spiniform tubercle at terminus. Areola narrow with one to two rows of punctations at narrowest point. Cervical spines absent; occasional faint singular tubercle present. Mandibular, branchiostegal and orbital regions of carapace with numerous small tubercles. Postorbital ridges short and ablated, rarely ending in a tubercle. Suborbital angle acute. Carapace cylindrical in profile. Antennal scale widest in middle, 1.7–3.1 (X ‾ = 2.5, n = 48, SD = 0.25) times as long as wide with slight undulation between distal and proximal ends. Spine of antennal scale curved outward at distolateral terminus. TCL 2.0–2.5 (X ‾= 2.2, n = 47, SD = 0.10) times longer than width. Form I and Form II males possessing hook on ischium of third periopods only. Hook on Form I males recurved gently, overarching basioichial joint; hook on Form II males weakly arched, never reaching basioichial joint. Hooks not opposed by tubercle on basis. Mesial surface of chelae with two rows of tightly spaced, staggered tubercles; mesial-most row with 5–7 (X ‾ = 6.4, n = 46, SD = 0.6) tubercles, second dorsal row with 3–5 (AAAAAA = 4.1, n = 46, SD = 0.4) tubercles, not extending above mesial surface of chelae onto body of palm. A single, subtle subpalmar tubercle occasionally present at dactyl junction with propodus. Single row of small punctations running along mesial longitudinal ridge of dactyl. Dorsomedian ridge of dactyl and opposable propodus pronounced. Ventral margin of propodus costate. Chelae normally lacking well-defined lateral impression at the junction of fixed finger with propodus base, occasionally stronger in larger individuals. Dactyl and propodus terminating in sharp corneous tip forming spine. One to two large carpal spines present with 3–4 carpal tubercles surrounding them. First pleopod of Form I male with short terminal elements. Central projection not tapering distally; recurved> 90° to main shaft of gonopod, with distinct subapical notch. Mesial process directed 90° to shaft deflected cephalolaterally; inflated cephalically, tapering to distinct caudal point beyond terminus of central projection. First pleopod of Form II male with central projection curved 90° to shaft, with complete apex; rounded. Mesial process not extending beyond central process, pointed and directed caudolaterally. Annulus ventralis, sclerotised, immovable and distinctly asymmetrical posteriorly, near rhomboid; cephalic portion with median trough leading to strongly sculptured central fossa; ‘S’ bend in sinus terminating at caudal edge.
Description of holotypic male, Form I
See Figure 4 View Figure 4 (a–k) and Table 3. Body cylindrical in shape ( Figure 4 View Figure 4 (h)); carapace posterior to cervical groove slightly wider than abdomen. Carapace width slightly greater than carapace depth at caudodorsal margin of cervical groove (12.3 and 11.8 mm, respectively, 95.8%). TCL 28.1 mm. Areola length 11.2 mm: areola 39.8% of TCL, 44.9% of postorbital carapace length (PCL). Rostrum moderately excavated; rostral margins thickened, slightly converging and continuous to base of acumen where margins become convergent and thin to tip. Rostrum floor with punctations. Rostrum 1.2 times longer than width at base. Acumen triangular terminating in upturned corneous tip ( Figure 4 View Figure 4 (h)). Postorbital ridges short, well developed, and lacking terminal spines or tubercles. Suborbital angle acute, lacking tubercle ( Figure 4 View Figure 4 (a)). Cervical spine absent. Mandibular, branchiostegal and orbital regions of carapace with numerous small tubercles; greatest tubercle density and definition in hepatic region.
Abdomen approximately equal in length to carapace, length 27.3 mm, width 10.2 mm. Pleura rounded cephaloventrally, angled distoventrally. Lateral margin of terga angulated. Lateral margin of second pleuron deeply furrowed. Cephalic section of telson with two large spines in each caudolateral corner. Proximal podomere of uropod with distomedian spine on mesial lobe. Mesial ramus of uropod with median ridge ending distally in distomedian spine just overreaching margin of ramus; laterodistal spine pronounced. Distal margin of proximal segment of lateral ramus of right uropod having 11 immovable, small spines and one large, lateral moveable spine. Cephalomedian lobe of epistome subcircular and cupped; zygoma moderately arched ( Figure 4 View Figure 4 (g)); cephalolateral margins thickened, forming sharp angle at junction with endostyle ( Figure 4 View Figure 4 (g)). Antennal scale broadest in middle, undulated along mesial length; lateral margin thickened, terminating in large corneous spine arcing outward at near 20° angle ( Figure 4 View Figure 4 (j)); mesial margin setiferous. Right antennal scale 3.8 mm long, 1.3 mm wide ( Figure 4 View Figure 4 (j)). Tip of right antenna reaching posterior portion of second pleuron when adpressed.
Mesial surface of right chelae palm with two rows of tubercles; mesial-most row with seven tubercles, second, more dorsal row with four small tubercles, smaller adpressed tubercles scattered lateral to second row; all tubercles with small setiferous tufts located on distal edge ( Figure 4 View Figure 4 (k)). Palm length (8.4 mm) 79.5% of palm width (10.6 mm); depth of palm 6.6 mm. Ventral surface of palm with a single subpalmar tubercle. Dorsal longitudinal ridge of dactyl developed and possessing multiple punctations with setiferous tufts ( Figure 4 View Figure 4 (k)). Dactyl and propodus terminating in large corneous spine. Dorsomedian ridge of fixed finger of propodus pronounced with punctations containing small setiferous tufts. No lateral impression. All measurements and counts from right chelae.
Carpus with prominent furrow and five weak dorsomedial tubercles; remainder of surface with some setiferous punctations; mesial margin with one large, procurved spine and three smaller spines near midlength. Distodorsal surface of merus with 10 weak tubercles; mesial-most ventrolateral ridge with 10 spines increasing in size distally; lateral-most ventrolateral ridge with seven small spines; ischium ventrolateral margin with three small spiniform tubercles and one large rounded tubercle. Hook on ischium of third periopods only; hook gently curved at apex, overarching basioischial joint, not opposed by tubercle on basis. Form I gonopod as described in diagnosis ( Figure 4 View Figure 4 (b–d)); tip reaching posterior margin of third caudomesial boss when abdomen is flexed.
Description of allotypic female
See Figure 4 View Figure 4 (i) and Table 3. Differing from holotype in the following respects: carapace depth slightly greater than carapace width (16.8 and 16.7 mm, respectively, 100.5%); TCL 36.9 mm, PCL 32.7 mm. Areola 40.0% of TCL (45.1% of PCL). Abdomen length 36.9 mm, abdomen width 13.3 mm. Mesial surface of chelae with two rows of tubercles; mesialmost row with six tubercles; second, more dorsal row with four tubercles. Palm length (10.6 mm) 85.9% of palm width (12.4 mm); depth of palm 7.6 mm. All measurements and counts from right chelae. Antennal scale 3.9 mm long, 1.6 mm wide. Annulus ventralis as described in diagnosis ( Figure 4 View Figure 4 (i)); first pleopods uniramous, reaching central region of annulus ventralis when abdomen flexed.
Description of morphotypic male, Form II
See Figure 4 View Figure 4 (e–f) and Table 3. Differing from the holotype in the following respects: Carapace depth less than carapace width (11.9 mm and 12.7 mm, respectively, 93.6%). Areola 40.6% of TCL (47.0% of PCL). Abdomen length 27.3 mm, abdomen width 9.9 mm.
Palm length (7.3 mm) 79.8% of palm width (9.2 mm). All measurements and counts from right chelae. Antennal scale 3.6 mm long, 1.3 mm wide. Gonopods reaching anterior margin of third periopod caudomesial boss. Central projection curved 90° to shaft, with complete apex; rounded ( Figure 4 View Figure 4 (f)). Mesial process does not extend beyond central process, pointed and directed caudolaterally. Hook on ischium of third pereiopod small, not reaching basioischial joint. Carapace scarred on left posterior side near junction with abdomen.
Colouration in life
Carapace, chelae and abdomen ground colour deep blue to cobalt ( Figure 3 View Figure 3 ); black colouration is a result of manganese buildup on the body due to heavy manganese deposits in the soils throughout the range of C. loughmani . Hepatic and antennal region punctuated with cream, blue, cobalt or white tubercles. Denticles on opposable surfaces of fingers cream, white or tan. Ventral surface of chelae white or cream fading to light blue on the lateral portions. Podomeres of pereiopods light blue, blue or cobalt; joints of pereiopodomeres cream or white. Ventral surface of abdomen and carapace cream or white. Dorsal ridge of Form I gonopod central projection amber; body of central projection, gonopod and mesial process tan. Form II gonopod and all associated processes cream to tan. Cephalic portion of annulus ventralis cream; caudal region of annulus ventralis ranges from cream to white. Ridge of fossa cream to tan.
Type locality
Wetland and ditch off Island Creek Road approximately 3.62 km (2.25 mi) northwest of Alum Creek, Lincoln County, West Virginia, USA (38.305497°N, − 81.853196°W; WGS84). All animals were excavated from their burrows. The wetland is fed by an intermittent headwater stream, originally connected to Island Creek before construction of the road. Canopy cover in the form of second-growth mixed mesophytic forest comprised of American sycamore ( Platanus occidentalis L.), eastern redbud ( Cercis canadensis L.), American basswood ( Tilia americana L.), sugar maple ( Acer saccharum Marshall ), black cherry ( Prunus serotina Ehrh. ), tulip poplar ( Liriodendron tulipifera L.) and spicebush ( Lindera benzoin L.). Herbaceous vegetation rooted within the wetland comprised of Japanese knotweed ( Fallopia japonica Houtt. ), wingstem ( Verbesina alternifolia L.), Canadian clearweed ( Pilea pumila L.), crooked stem aster ( Symphyotrichum prenanthoides ((Muhl. ex Willd.) G. L. Nesom)), swamp buttercup ( Ranunculus hispidus Michx. ), Japanese stiltgrass ( Microstegium vimineum (Trin.) A. Camus ), jewelweed ( Impatiens capensis Meerb. ), cinnamon fern ( Osmunda cinnamomea L.) and fowl bluegrass ( Poa palustris L.). Soils at the site characterised by dark sandy silt loam in the upper 3–6 cm, followed by a thick clay layer below with low-chroma soils typically interspersed with manganese masses. The type series was collected during the months of July and October with David A. Foltz II collecting the allotype and morphotype on 8 July 2017 and David A. Foltz II and Greg A. Myers collecting the holotype on 14 October 2017.
Disposition of types
The holotype, allotype and morphotype of C. loughmani are deposited in the North Carolina Museum of Natural Sciences ( NCMNS), Raleigh, NC (catalogue numbers 90082, 90083, 90084). Paratypes are deposited in the National Museum of Natural History ( USNM), Smithsonian Institution, Washington, DC (catalogue number USNM 1480610 View Materials ), Carnegie Museum of Natural History ( CMNH), Pittsburgh , Pennsylvania (catalogue number CMNH 38,828 View Materials ), and the Ohio State University Museum ( OSUM), Columbus , Ohio (catalogue number OSUM 10500 ) .
Geographic range and specimens examined
Cambarus loughmani is a West Virginia endemic, restricted to a 3265 km 2 (1261 mi 2) area of the Appalachian Plateau physiographic province in Cabell, Kanawha, Lincoln, Mason, and Putnam counties ( Figure 5 View Figure 5 ) . It is endemic to the pre-glacial Teays River Valley, named for the Teays River which once flowed through the area, conveying drainages now associated with the upper Ohio River through northern Ohio, Indiana and Illinois . Its currently understood range is limited to this region by the Ohio, Kanawha, Coal and Mud rivers which function as boundaries between it and the RCBB morph . The majority of its range falls north of the pre-glacial Teays River Valley, through which the Mud River now flows . Collecting efforts were conducted throughout this range; however, much of the lowland habitat, particularly that through the Teays Valley and floodplains along the major rivers, has been converted to agricultural, residential or industrial use . Access to the lower portion between the Mud and Coal river headwaters was impossible to access, as a large surface mine covered much of the area .
During the study, an image of C. loughmani was uploaded to a species identification application named iNaturalist by an employee at Chief Logan State Park located in Logan County, West Virginia. We investigated the area and found a large population of C. loughmani in the park; however, the area is 35 miles to the southwest of our southernmost site for the species and falls outside of the boundaries for its range. Further investigations outside the park yielded the RCBB morph in the first small watersheds immediately north, south, east and west of the site. Upon closer inspection, we determined the wildlife area of the park had had a substantial amount of earth brought in to raise and level the area and to construct a dam within the park to establish a lake for fishing. These events took place around 1996, during a time of substantial economic and housing growth within the Teays Valley and I-64 corridor. While records of exactly where the fill originated could not be found, it is not out of the realm of possibility that the animals were brought in with the fill and became established following the covering and compaction of the original ground. This is further substantiated by finding the RCBB morph in watersheds bracketing the park and the fact that Lacunicambarus thomai ( Jezerinac, 1993) (previously Cambarus thomai ; see Glon et al. 2018), another burrowing species, has previously been reported as introduced into the Deep Creek Lake region of Maryland ( Loughman 2010). We therefore view the population of C. loughmani located within Chief Logan State Park as introduced.
Due to its limited distribution, and potential conservation concerns associated with over collecting, specific locations coordinates will not be given following collection accession numbers, but are available from the primary author upon request. A total of 118 specimens of C. loughmani were examined from 25 localities which comprise the following 33 lots:
WEST VIRGINIA: Cabell County: 20170914-01, pond near Ona, WV, S of I-64, 14 September 2017, 2♂ I 4♀, Timothy Brust ( TB); WSWV, Spurlock Creek of Guyan Creek of Ohio River at Union Ridge Road, 3.8 mi . E of Green Bottom , 1.75 mi . N of Union Ridge , 3 May 1987, 4♀, 2♀ j ., 3♂ YOY, 2♀ YOY, Raymond F. Jezerinac ( RFJ), Greg Lombardo ( GL), Randy French ( RF), Dave Chrisman ( DC), G . Whitney Stocker ( GWS); J-87-78 , Trace Fork of Guyan Creek of Ohio River, 2.0 mi . N of Dudley Gap , 3.6 mi . N of Milton 5.5 mi . SE of Glenwood , 12 Sept . 1978, 3♂ I, 2♀, GWS, DC, Roger F . Thoma ( RFT), Zachary B . Thoma ( ZBT), RFJ; J-87-77 , intersection of Co . Rd. 1 and Co. Rd. 1/1, 0.5 mi . W of Gwinn , 12 Sept . 1987 , 1♀, GWS, RFJ, DC, RFT, ZBT; J-89-38 , intersection of Co. Rd. 12 and Co. Rd. 7, 2.3 mi . NNE of Lesage , 14 June 1989, 1♂ I, 1♀, GWS, RFJ;. Kanawha County: J-89-40 , County Road 3, 1.61 km SW of CR9 [1.77 km SW of Tornado], 21 June 1989, 1♀, GWS and RFJ . Lincoln County: 20170708-3, road pull off near creek 3.62 km NW of Alum Creek along Island Creek , 7 July 2017, 1♂ II 4♀, David A . Foltz II ( DAF); 20170819-5, road pull off near creek 3.62 km NW of Alum Creek along Island Creek, 19 August 2017, 1♂ I 2♀ 1 juv ., DAF and Ashley J . Leek ; 20171014-3, road pull off near creek 3.62 km NW of Alum Creek along Island Creek, 14 October 2017, 1♂ I 1♀ ., DAF and Greg Myers ( GM) . Logan County: 170821-01, seep at Chief Logan State Park next to wildlife center, 21 August 2017, 1♂ I, GM; 201710-13-01, pull off at security gate leading to Chief Logan Wildlife Center, 13 October 2017, 2♀, DAF and Nicole M . Sadecky ( NMS); 20171013-1, pull off at security gate leading to Chief Logan Wildlife Center, 13 October 2017, 1♂ I 6♀, DAF . Mason : 100601-11-ZL, Chief Cornstalk Hunting Grounds Campground [7.52 km SW of Beech Hill], 1 June 2010, 2♀, DAF and Zachary J . Loughman ( ZJL); 170622-01, roadside ditch along Keister Road, 22 Jun 2017, 3♀, GM; 170623-04, roadside ditch along Rocky Fork Road 30.5 m upstream of bridge, 23 June 2017, 1♂ I 2♂ II 4♀ 1 juv ., GM; 20170519-01, house on Spanoaks Drive 1.61 km S of Upton, 19 May 2017, 1♀, ZJL and Mael G . Glon ( MGG); 20170617-2, residential yard ditch on Ashton-Upton Road 3.1 km S of Mt . Olive , 17 June 2017, 1♂ I, David A . Foltz II; 20170617-7, Chief Cornstalk Hunting Grounds Campground [7.52 km SW of Beech Hill], 17 June 2017, 1♀, DAF; J-86-30 , Chief Cornstalk Hunting Grounds Campground [7.52 km SW of Beech Hill], 20 April 1986, 1♀, GWS, RFJ, and DC; J- 87-79 , CR 86, just W of CR 43 [3 km SW of Mount Olive, 12 September 1987, 3♀, GWS, RFJ, DC, ZBT, and RFT; WSWV-8618 , Chief Cornstalk Hunting Grounds Campground [7.52 km SW of Beech Hill], 31 May 1986, 1♀, GWS; WSWV-8701 , Chief Cornstalk Hunting Grounds Campground [7.52 km SW of Beech Hill], 2 May 1986, 1♀, GL, RF, and GWS; WSWV-8702 , CR 27/3, 0.8 km W of CR 27 [4.35 km NE of Arlee], 2 May 1987, 4♀, GL, RF, and GWS . Putnam : 20170519-02, hillside seep along road 1.93 km due E of Upland, 19 May 2017, 1♂ I 1♂ II 1♀, ZJL and MGG; 20170519-03, CR 13-8 approximately . 64 km NE of intersection with CR-40 [6.44 km NW of Teays Valley], 19 May 2017, 2♂ II, ZJL and MGG; 20170708-4, 0.8 km N of Teays Valley, 8 July 2017, 6♀, DAF; 20171009-1, Poindexter Branch Seep , 5.31 km N of Hurricane and I-64, 9 October 2017, 2♂ I 3♀, TB; J-87-80 , CR 29/5, 0.32 km NW of CR 29 [7.72 km NNW of Nitro], 13 September 2017, 1♀, GWS, RFJ, DC, ZBT, and RFT; J-88-308 , Hwy 60, 0.32 km W of CR 60/18 [4.67 km SW of Teays], 1 October 1988, 1♂ I 1♀, RFJ, GWS, and Tom Jones (TJ); J-88-309 , Co Rd 37, 2.74 km S of CR 39 [10.46 km S of Hurricane], 1 October 1988, 1♀, RFJ, GWS, and TJ; TJ-89-04 , CR-32, 2.41 km W of WV Rt . 34, 2.01 km NW of Mt . Vernon , 2.82 km N of Teays Valley, 1 May 1989, 4♂ I 6♂ II 10♀ 1 juv ., TJ; 170710-1, seep along Bias Hill Road , 10 July 2017, 1 juv ., NMS and DW Mahan ( DWM); 170730-1, seep along Bias Hill Road, 30 July 2017, 1 juv ., NMS and DWM.
Conservation status
Cambarus loughmani should be listed as Threatened using the American Fisheries Society criteria ( Taylor et al. 2007, 376): ‘a species likely to become endangered throughout all or a significant portion of its range’. Because of its narrow range the loss of only a few populations to activities such as draining of wet areas, an activity undertaken by most homeowners, would result in significant losses. It should be assigned a G3 (Vulnerable = ‘At moderate risk of extinction due to a restricted range, relatively few populations (often 80 or fewer), recent and widespread declines, or other factors’) ranking using the Global Conservation Criteria ( Masters 1991, 560) and Endangered B1b(i,iv) using IUCN (2001) Red List criteria.
Ecology and life history
Cambarus loughmani sp. nov. is best classified as a primary burrower ( Hobbs 1981) as it utilises burrows for all known aspects of its life history. Although some colonies were located adjacent to surface waters, no animals were observed in surface waters. The species appears to prefer wet meadows, seepages, wet ditches and other areas with perched water tables. Cambarus loughmani is most readily found in wet roadside ditches or seepages with little to no connectivity to other surface waters, and usually at the crest of hills throughout its range. It remains unclear whether these areas are preferred or merely unutilised by other burrowing species within C. loughmani ’s range that might be competitors. While C. loughmani can be found with other burrowers, its presence within a colony is usually less dense than that of heterospecifics, particularly when found alongside L. thomai . At CCWMA, C. loughmani inhabits the wet field near the lake along with L. thomai ; however, L. thomai is encountered more often near the lake, while C. loughmani is encountered more readily farther away from the lake (DAF, personal observation). Burrowing colonies comprised entirely of C. loughmani are rare and are more commonly found on the crests of hills where no connectivity to another water source is present.
Burrow morphology for C. loughmani in natural to near-natural conditions is typically simplistic, consisting of burrows with two entrances in a basic ‘Y’ shape for 10–30 cm leading to a vertical shaft 25–60 cm long. After the primary shaft, a small chamber is typically present near or below the water table which continues to and terminates in a resting chamber. The main entrance typically consists of a chimney while the secondary entrance is often flush with or countersunk into the ground and covered with vegetation or leaf litter. Resting chambers often contained vegetation such as fallen leaves, fleshy stalks, seeds, roots and small pieces of wood. Ancillary tunnels from the central shaft or resting chamber were rare and only noted in areas with high burrow density (> 10/m 2). In one instance, three separate shafts ran to a large voided chamber approximately 30 cm below the surface. Unlike the typical simplistic resting chambers utilised by the species, these shafts continued downward an additional 20 cm before terminating in another resting chamber. One chamber was empty; however, the other two each contained a female C. loughmani , indicating multiple animals utilising a connected burrow system.
While C. loughmani typically followed the aforementioned simplistic, relatively shallow burrow design, areas with deep water tables such as pastures and heavily downcut stream banks at Chief Logan State Park contained single-shaft burrows ranging from 90 to 220 cm before reaching the water table, followed by another 20–30 cm to the resting chamber.
Soils that C. loughmani can be found in throughout its range are variable. Unaltered to nearly unaltered areas often consist of at least two to four soil horizons, with the top layers usually consisting of some form of sandy to loamy dark soil while lower soil layers are typically clay to clay-loam and hydric, indicated by low chroma, mottling, and manganese masses found throughout the region. Sandstone fragments and trapped organics may also be present. In highly altered areas, the general soil composition is comparable; however, it is usually characterised by reduced soil horizons, heavy amounts of trapped organics, and trapped or mixed particles of rock or other materials atypical of the region or soil surface, including metal, glass, obsidian, slag, brick, coal and other materials.
Vegetation rooted in and around the burrows of C. loughmani in unaltered to nearly unaltered areas is also variable, but is typically comprised of the plants listed in the description of the type locality, with occasional shifts in community typical of a mixed mesophytic forest within the physiographic area. In altered areas, however, such as fields and yards, the vegetation was drastically different and included white clover ( Trifolium repens L.), red clover ( Trifolium pretense L.), timothy ( Phleum pretense L.), common plantain ( Plantago major L.), red fescue ( Festuca rubra L. ssp. rubra ), orchard grass ( Dactylis glomerata L.), Queen Anne’ s lace ( Daucus carota L.), horse nettle ( Solanum carolinense L.) and other plants commonly associated with upland farm fields within the area. Despite plant densities and community, C. loughmani appears to be indiscriminant in the vegetative portion of its diet, as plants and leaves within the burrows often consisted of whichever plants were nearest and most accessible to the burrow entrance. Finding entire herbaceous plants with snipped or grazed leaves and upper portions pulled completely into burrows was common.
The life history of C. loughmani is poorly understood at this time and warrants further investigation to better elucidate our understanding of the species. During collections in June, a large Form I male showing signs of pre-moult was collected from an excavated capped burrow in a roadside ditch in Mason County, West Virginia. This male was heavily laden with manganese buildup on its carapace to the point that it appeared almost black. During excavation efforts, part of the animal’ s telson was damaged, revealing a soft, bright blue telson unlike the hardened portion of the exoskeleton, seemingly confirming the animal was about to undergo ecdysis.
Collections of C. loughmani during the months of June through October were attempted using both baited lines and excavation, with excavation serving as the primary method for delineating the animal’ s range while baited lines served as the primary method for collecting large numbers of animals quickly when colonies were found. During June, following a heat wave, only one C. loughmani was observed and collected through the use of a baited line. This single female was actually in the process of removing clay from its burrow, presumably in response to water table draw-down. Despite the lack of feeding behaviour, baited line was attempted and the animal immediately dropped the clay pellet it was carrying and pursued the bait when it touched its chelae. This behaviour highlights the strong feeding response of the animal. During the month of July, following heavy rains, at night C. loughmani was observed at the surface more frequently. Burrow colonies at this time consisted of a mix of capped and uncapped burrows. The uncapped burrows housed females, sitting at the entrance of their burrows at night, while the capped burrows contained Form II males. Both of these observations were confirmed through excavation of burrows. During collections in August and September, few to no capped burrows were witnessed at sites and Form I males were present and active alongside females in the evenings. During collections in October, both males and females were present at the portals to their burrows at night; however, at later points in the month, C. loughmani were observed pulling leaves into and plugging their burrows. Additionally, during mid-October a Form I male was observed on the surface traversing from burrow to burrow. It encountered two females, both of which came out towards the male with claws agape. Following a brief display of touching each other’ s antennae and swinging their chelae back and forth in slashing motions, by both the male and the female, the female retreated to the portal of her burrow and positioned her chelae agape so as to block the entrance. The male continued onward to another burrow, and was collected after the second observance of this behaviour.
In colonies where C. loughmani occurred with heterospecifics, we noted that it was the last species to rise to the surface at night. Both L. thomai and C. b. cavatus were present at their burrow portals shortly after twilight, but C. loughmani typically did not rise to the surface until a few hours after twilight.
Crayfish associates
Lacuni cambarus thomai and Cambarus b. cavatus Hay, 1902 have been collected from burrows directly alongside C. loughmani in suitable habitats. Although not yet collected alongside C. loughmani , Creaserinus fodiens ( Cottle 1863) and Procambarus acutus Fowler 1912 are found in the lowland wetlands of its range, with C. fodiens being restricted to relict Acer dominated floodplains along the Ohio River and P. acutus restricted to swamps and backwaters along the Kanawha River ( Loughman and Simon 2011). Faxonius sanbornii ( Faxon 1884) can be found in flowing streams throughout the area C. loughmani is known from, and non-native Faxonius virilis ( Hagen 1870) has been collected from ponds within CCWMA less than 15.2 m (50 ft) from C. loughmani burrows. Cambarus theepiensis Loughman et al. 2013a inhabits large flowing waters in the Guyandotte and Mud rivers, while Cambarus robustus Girard 1852 is found in the same habitat within the Kanawha and Coal Rivers. Lastly, in the southernmost portion of its range, the RCBB morph of C. aff. dubius can be found; however, to date no C. loughmani have been collected alongside C. aff. dubius .
Relationships and comparisons
Prior to Crandall and De Grave (2017), C. loughmani sp. nov. would have been placed within the subgenus Jugicambarus , along with other members of the C. dubius complex, based on the presence of a row of cristiform tubercles located on the mesial margin of their chelae ( Hobbs 1969). However, Crandall and De Grave (2017) revised the taxonomy for North American crayfishes in the genus Cambarus , removing the subgenera due to lack of phylogenetic validity. While they state some groupings of the subgenera may be valid and warrant elevation to genus level, we present no such data for elevation of Jugicambarus herein and will, instead, refer to C. loughmani ’s similarities to other members of the C. dubius complex from which it is described. In particular, it bears a striking resemblance to C. pauleyi based on its blue colouration in life, and two rows of tubercles located along the mesial margin of the palm of the chelae. In fact, Jezerinac et al. (1995) previously grouped C. loughmani and C. pauleyi together as ‘blue morph dubius’; however, upon closer inspection, the two can be differentiated readily by comparison of the areola, where in C. pauleyi it is obliterated and in C. loughmani it is open and contains one to two rows of punctations at its narrowest point. Additionally, C. pauleyi has one (occasionally two) pronounced subpalmar tubercles while C. loughmani has either zero or one subtle subpalmar tubercle. Lastly, the lateral edge of the antennal scale in C. pauleyi forms a straight line from the most distal to proximal points, while in C. loughmani the distolateral edge arcs out away from the proximal base of the scale. Other species of the C. dubius complex are easier to differentiate from C. loughmani based on colour in life and morphological features. Cambarus adustus is brown compared to the deep blue of C. loughmani , while C. dubius is bright orange to red. Both C. adustus and C. dubius also have a single row of cristiform tubercles on the mesial margin of the chelae, while C. loughmani has two tight, well-defined rows.
Additionally, as the C. dubius complex is widespread, difficult to collect, and understudied, it is worth noting comparisons of other regional morphs that have yet to receive treatment, but could be confused with C. loughmani . The first of these, the nearby C. aff. dubius , was treated by Jezerinac et al. as the RCBB morph. In life, C. loughmani can be differentiated from this morph by comparison of colour as the RCBB morph is exactly as it sounds, occasionally with subtle shifts in brightness and location of the colours, but always with some form of red-orange claws and blackish body while C. loughmani is always blue. Additionally, nearby populations of the RCBB morph have only a single row of tubercles on the mesial margin of the chelae palm, as opposed to two rows in C. loughmani .
Etymology
The authors of this paper name this crayfish in honour of Dr Zachary J. Loughman, assistant professor of biology, West Liberty University. In recent years, prior to the naming of this species, Dr Loughman has served as one of the primary astacological researchers, contributing greatly to our understanding of ecology, taxonomy, conservation and distribution of Appalachian crayfish species. It is fitting that this crayfish be named in his honour, as both he and this crayfish are reclusive, hard to track down, and when faced with adversity never back down and often advance with arms flailing. The authors of this paper are composed of a mix of age classes. For the younger authors, Dr Loughman has served as a teacher, advisor and mentor. For the older authors he has served as a colleague, collaborator and student. For all of us, he has been a friend and an inspiration.
Common name
The suggested common name for this species is blue Teays mudbug as it inhabits the Teays River Valley.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cambarus loughmani Foltz, Sadecky, Fetzner and Thoma
| Foltz, David A., Sadecky, Nicole M., Myers, Greg A., Fetzner, James W., Welsh, Stuart A., Stocker, G. Whitney, Glon, Mael G. & Thoma, Roger F. 2019 |
Cambarus aff. dubius
| Loughman ZJ & Thoma RF & Fetzner JW Jr & Stocker GW 2015: 534 |
Cambarus (Jugicambarus) dubius
| Jezerinac RF & Stocker GW & Tarter DC 1995: 121 |
| Hobbs HH Jr. 1989: 22 |
| Hobbs HH Jr. 1974: 18 |
| Hobbs HH Jr & Bouchard RW 1973: 62 |
Cambarus carolinus dubius
| Faxon W 1914: 396 |
Cambarus dubius
| Dewees JP 1972: 1 |
| Faxon W 1885: 70 |
| Faxon W 1884: 114 |