
Veromessor chamberlini (Wheeler, 1915)
|
publication ID |
https://doi.org/10.11646/zootaxa.5206.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CE749F6C-5832-4152-AB4B-6D89ACCDD560 |
|
DOI |
https://doi.org/10.5281/zenodo.7327869 |
|
persistent identifier |
https://treatment.plazi.org/id/039387FD-FFFF-FFD4-7FC5-FF1B58CAF839 |
|
treatment provided by |
Plazi |
|
scientific name |
Veromessor chamberlini |
| status |
|
( Figures 1B View FIGURE 1 , 6C View FIGURE 6 , 11 View FIGURE 11 , 12A View FIGURE 12 , 13–14 View FIGURE 13 View FIGURE 14 )
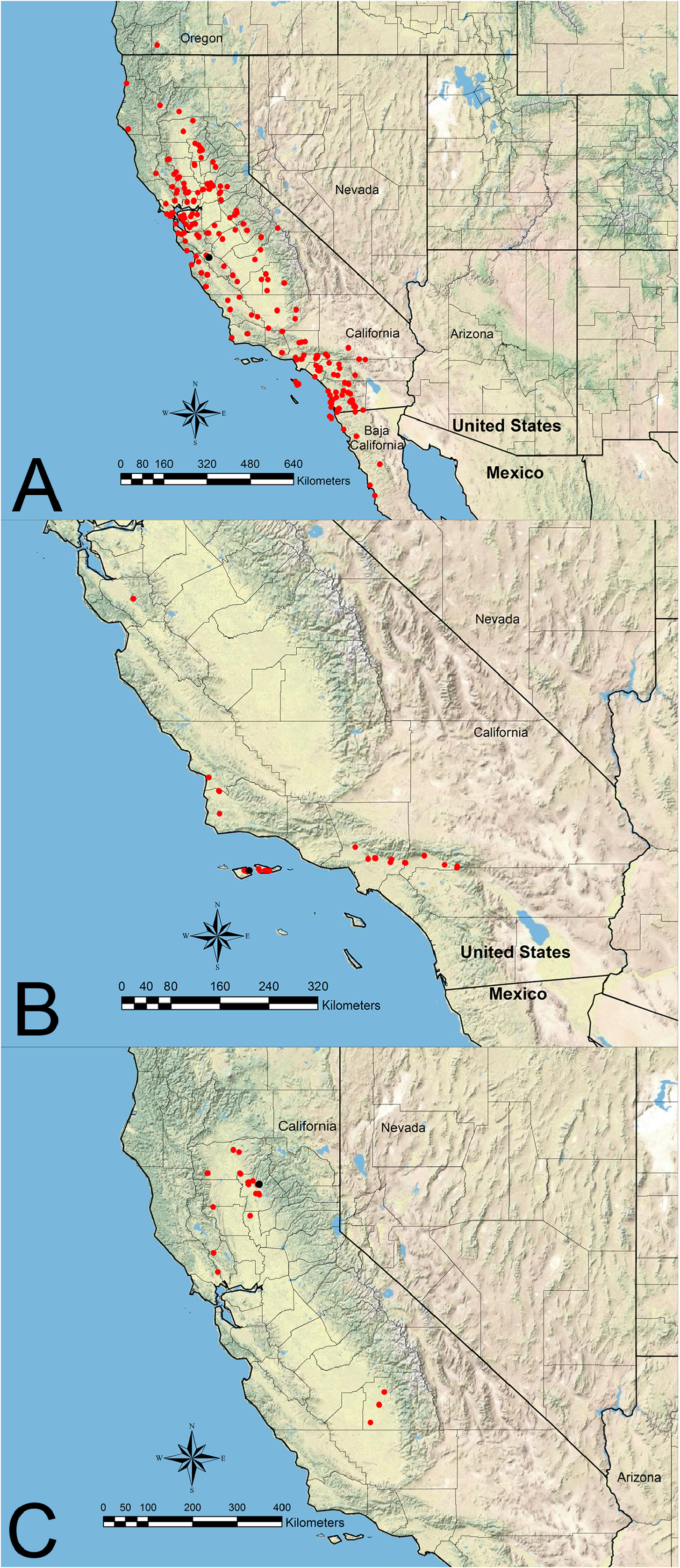
Distribution— Figure 10B View FIGURE 10
Messor chamberlini Wheeler, 1915: 410 View in CoL (worker). Syntypes examined: 3 workers [ LACM], 6 workers [ MCZC], 3 workers [USNM], UNITED STATES, California, Santa Barbara County, Santa Cruz Island, no date (Dr. R.V. Chamberlin); Wheeler and Wheeler, 1972: 240 (larvae); Enzmann, 1947: 152 (as subspecies of Veromessor andrei ); Creighton, 1950: 159 (revived status as full species). See also Wheeler and Creighton, 1934: 366, plate II, fig. 5. USNM worker here designated LECTOTYPE [USNMENT00529080].
Novomessor ( Veromessor) chamberlini (Wheeler) ; Emery, 1921: 67 (first combination in Novomessor [ Veromessor ]).
Veromessor chamberlini (Wheeler) ; Wheeler and Creighton, 1934: 366 (first combination in Veromessor ).
Novomessor ( Veromessor) andrei subsp. chamberlini (Wheeler) ; Enzmann, 1947: 152 (revived combination as subgenus of Novomessor and first combination as subspecies of andrei ).
Veromessor chamberlini (Wheeler) ; Creighton, 1950: 157 (revived combination in Veromessor ; re-elevated to specific status).
Messor chamberlini Wheeler View in CoL ; Bolton, 1982: 341 (revived combination in Messor View in CoL ).
Veromessor chamberlini (Wheeler) ; Ward, Brady, Fisher & Schultz, 2015: 13 (revived combination in Veromessor ).
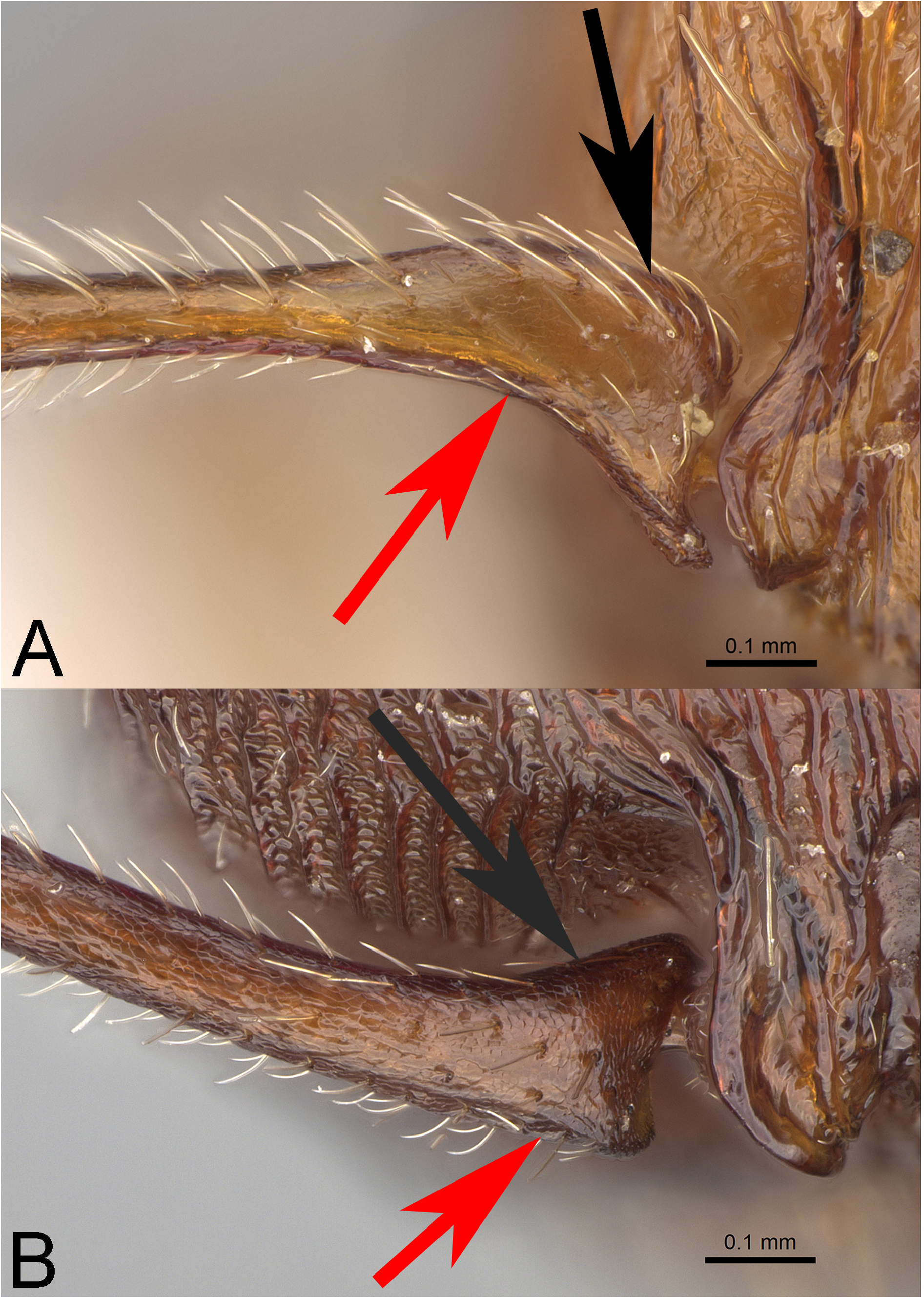
Worker diagnosis. This species is uniquely characterized by the following combination of features: (1) yellowishred to dark orangish-red, (2) medial lobe of clypeus with two strong submedial rugae, sometimes with one to two weaker sublateral rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4) dorsal base of scape flattened and widened; maximum basal width of scape distinctly greater than maximal preapical width; in frontal view, base of scape asymmetrical with ventral portion of anterior edge greatly expanded compared to dorsal edge, (5) MOD less (usually much less) than OMD, OI <22.0, (6) cephalic dorsum with prominent wavy to weakly irregular rugae, medial rugae diverging toward posterior corners; rugae along posterior margin strongly irregular to weakly rugoreticulate; cephalic interrugae weakly to moderately coriarious, moderately shining, (7) psammophore moderately well developed; ventral surface of head capsule with J-shaped hairs arranged in a V-shaped row which does not reach the posterior part of lateroventral margin of head capsule, usually mixed with straight or evenly curved hairs, (8) dorsum of pronotum with weakly to strongly irregular rugae; sides of pronotum with widely spaced, regular to wavy, longitudinal rugae; mesonotum with irregular longitudinal rugae; mesopleura with regular to wavy rugae that traverse longitudinally to posterodorsally; interrugae on mesosoma weakly coriarious, shining, (9) propodeal spines slender, acuminate, weakly curved posterad in profile and weakly curved inward in dorsal view; length> 3.0× the distance between their bases and length> 1.0× MOD; infraspinal facet and propodeal declivity weakly coriarious to weakly rugose, strongly shining, and (10) metasternal process moderately large, longer than high, apex broadly rounded, margin irregular and translucent ( Figures 6C View FIGURE 6 , 11 View FIGURE 11 , 12A View FIGURE 12 ).
Measurements. lectotype ( n = 32 + 1 paralectotype). HL 1.32 (1.11–1.45); HW 1.33 (1.06–1.42); MOD 0.24 (0.21–0.27); OMD 0.34 (0.23–0.39); SL 1.06 (0.92–1.15); PNW 0.83 (0.67–0.90); HFL 1.39 (1.14–1.53); ML 1.68 (1.33–1.88); PW 0.28 (0.20–0.29); PPW 0.43 (0.34–0.46). Indices: SI 79.70 (73.23–87.74); CI 100.76 (94.12– 104.17); OI 18.05 (16.30–21.70); HFI 104.51 (93.60–110.16).
Queen diagnosis. This caste is diagnosed by the following combination of features: (1) orangish to orangishbrown, (2) medial lobe of clypeus with two strong submedial rugae, sometimes with one to two weaker sublateral rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4) dorsal base of scape flattened and widened; maximum basal width of scape distinctly greater than maximal preapical width; in frontal view, base of scape asymmetrical with ventral portion of anterior edge greatly expanded compared to dorsal edge, (5) MOD similar to OMD, (6) cephalic dorsum with prominent wavy to weakly irregular rugae, medial rugae diverging toward posterior corners; rugae along posterior margin weakly irregular to weakly rugoreticulate; interrugae on cephalic dorsum weakly to moderately coriarious, weakly shining, (7) psammophore moderately well developed, (8) mesoscutum shining with fine longitudinal rugae, midline broadly smooth and shining between fine longitudinal rugae, (9) propodeal spines shorter than distance between their bases; infraspinal facet smooth and shining, and (10) metasternal process moderately large, longer than high, apex broadly rounded, margin irregular and translucent ( Figure 13 View FIGURE 13 ).
Measurements. ( n = 4). HL 1.42–1.47; HW 1.44–1.50; MOD 0.33–0.36; OMD 0.35–0.36; SL 1.12–1.25; HFL 1.52–1.61; ML 2.30–2.49; PW 0.37–0.41; PPW 0.58–0.61. Indices: SI 77.24–83.33; CI 99.31–103.52; OI 22.76–24.49; HFI 101.33–111.81.
Male diagnosis. This caste is diagnosed by the following combination of features: (11) blackish brown, (12) *, (13) *, (14) *, (15) anepisternum densely lineopunctate; katepisternum strongly coriarious, anterior one-half of katepisternum with scattered, long, flexuous hairs, posterior one-half lacking long flexuous hairs except for a row of short hairs along posterior border, (16) propodeum with widely spaced longitudinal rugae, interrugae densely punctate, dull; spines absent, (17) metasternal process triangular, acute, with lamella extending on each side, portion posterior to lamella longer than portion anterior to lamella, and (18) subpetiolar process bluntly triangular, apex acute ( Figures 1B View FIGURE 1 , 14 View FIGURE 14 ). * = head missing.
Measurements. ( n = 1). HL *; HW *; MOD *; OMD *; SL * HFL 1.68; ML 2.09; PW 0.27; PPW 0.48; AOD *; IOD *; OOD *. Indices: SI *; CI *; OI *; HFI *. * = head missing.
Additional material examined. UNITED STATES: California: Los Angeles Co.: Pasadena, Nov 11, 1922 & May 13, 1946 & June 13, 1946 (LACM; UCDC; USNM); San Gabriel River at Irwindale, Feb 17, 1963 & May 9, 1963 & Jun 19, 1963 & Feb 22, 1965 (CASC; LACM; RAJC; UAIC; UCDC; USNM); Azusa, Dec 5, 1928 & Jun 14, 1931 (USNM); Claremont, Nov 16, 1945 (UCDC; USNM); 2 mi E San Fernando, May 7, 1931 (USNM); Glendale, July 1941 (UCDC). San Bernardino Co.: Mentone, Aug 22, 1965 (LACM); Oak Glen, 1500 m, Sep 23, 1982 & May 2, 1983 & Jul 26–Aug 2, 1983 & Jul 1–14, 1985 & Aug 1, 1985 (CASC; LACM; UCRC); Rialto at Riverside & Sierra Avenues, 1870’, Jul 20, 1992 (RAJC). San Luis Opisbo Co.: Arroyo Grande, Aug 2–3, 1988 (UCDC). Santa Barbara Co.: Santa Maria (as San Maria on one label), Jun 8, 1957 & Aug 6, 1957 & Aug 29, 1957 (LACM; USNM); Santa Cruz Island at E end of central valley, May 23, 2013 (RAJC); Santa Cruz Island at Field Station, 70 m, Jun 26, 1993 & Jul 4, 2003 & Aug 4, 2003 (NHMW; RAJC; UCDC); Santa Cruz Island at South Ridge, 430 m, Jun 27, 1993 (RAJC); Santa Rosa Island at Cherry Canyon, 45 m, Apr 12, 2014 (MLBC; UCDC); Santa Rosa Island at Windmill Canyon, 45 & 85 m, Apr 12–14, 2014 (UCDC); Santa Cruz Island, Apr 18, 1936 (USNM); Santa Cruz Island at El Tigre Ridge, May 4, 2000 (UCDC); Santa Cruz Island at Torrey Pines, 30 m, Apr 13, 2014 (MMPC; UCDC); La Purisima Mission in NE Lompoc, 60 m, May 25, 1985 (JTLC); La Purisima at 5 km NE Lompoc, 125 m, Jun 12, 1994 (record from M. Bennett, pers. comm.). Santa Clara Co. : no loc, no date (LACM). Questionable locales (appear to be or are outside of geographical range): UNITED STATES: Missouri: Barton Co.: no loc, no date (MCZC) ( Figure 10B View FIGURE 10 ).
Etymology. This species was named to honor Dr. D.V. Chamberlin, who collected the type series on Santa Cruz Island.
Discussion. Veromessor chamberlini appears to have a relatively small geographic range, such that it might be sympatric only with V. andrei and V. stoddardi . Workers of V. chamberlini are separated from both species based on: (1) moderately well developed psammophore, and (2) posterior margin of frontal lobes notably constricted. Both V. andrei and V. stoddardi have: (1) poorly developed psammophore, and (2) posterior margin of frontal lobes lacking a constriction.
A molecular phylogeny that used UCEs shows V. chamberlini and V. andrei are sister lineages (M.L Borowiec, unpub. data).
Biology. Little is known about the biology of V. chamberlini . Workers are diurnal and forage in a column (M. Bennett, pers. comm.). Colonies are small with a maximum of 627 workers (M. Bennett, pers. comm.). Workers are monomorphic.
Gland chemistry has not been examined in V. chamberlini . Like other small-colony congeners, workers of V. chamberlini have a small pygidial gland reservoir and lack a textured tergal cuticle ( Hölldobler et al., 2013).
Mating flights have not been observed, but sexuals have been collected in Malaise traps from July 1 through Aug 2, indicating that mating flights occur during summer.
The geographic range of V. chamberlini appears to be restricted to coastal islands and adjacent mainland coastal areas of southern California and extending inland along the southern base of the San Gabriel Mountains. Most potential habitat south of the San Gabriel Mountains has been developed. There also is one disjunct record from Santa Clara County that did not indicate a collector, locale, or date. The veracity of this record is unknown, but it seems likely that the historical range of V. chamberlini extended along coastal areas north to or near the San Francisco Bay area. Efforts should be made to reverify this northern record. This species occurs at elevations from 30–1,500 m in the California interior chaparral and woodlands, California coastal sage and chaparral, and California montane chaparral and woodlands ecoregions, as defined by Olson et al. (2001) ( Figure 10B View FIGURE 10 ).





| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Myrmicinae |
|
Genus |
Veromessor chamberlini
| Johnson, Robert A., Borowiec, Marek L., Snelling, Roy R. & Cole, Arthur C. 2022 |
Veromessor chamberlini (Wheeler)
| Ward, P. S. & Brady, S. G. & Fisher, B. L. & Schultz, T. R. 2015: 13 |
Messor chamberlini
| Bolton, B. 1982: 341 |
Veromessor chamberlini (Wheeler)
| Creighton, W. S. 1950: 157 |
Messor chamberlini
| Wheeler, W. M. 1915: 410 |