
Veromessor stoddardi (Emery, 1895)
|
publication ID |
https://doi.org/10.11646/zootaxa.5206.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CE749F6C-5832-4152-AB4B-6D89ACCDD560 |
|
DOI |
https://doi.org/10.5281/zenodo.7327877 |
|
persistent identifier |
https://treatment.plazi.org/id/039387FD-FFBE-FFAD-7FC5-FF1B5FD7FEED |
|
treatment provided by |
Plazi |
|
scientific name |
Veromessor stoddardi |
| status |
|
( Figures 1J View FIGURE 1 , 48–51 View FIGURE 48 View FIGURE 49 View FIGURE 50 View FIGURE 51 )
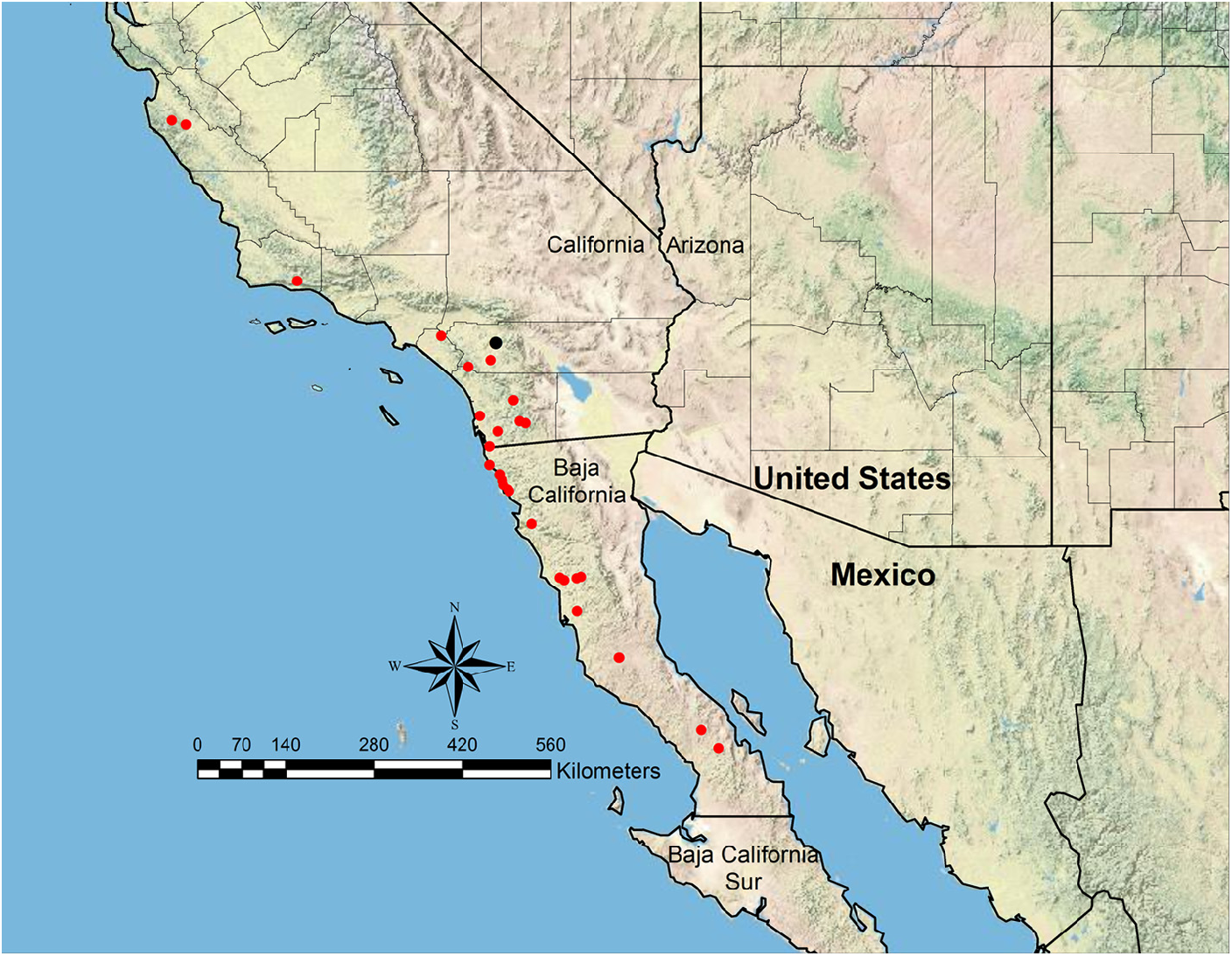
Distribution— Figure 52 View FIGURE 52
Stenamma ( Messor) stoddardi Emery, 1895: 307 (worker). Syntypes examined: 2 workers [ LACM], 7 workers [ USNM] , #55466, UNITED STATES, California: Riverside County, San Jacinto (Hyatt?). See also Wheeler and Creighton 1934: 385, plate II, fig. 1. USNM worker here designated LECTOTYPE [USNMENT00922798].
Novomessor stoddardi (Emery) ; Emery, 1915: 73 (first combination in Novomessor ).
Novomessor ( Veromessor) stoddardi (Emery) ; Forel, 1917: 235 (first combination in Novomessor ([ Veromessor ]).
Veromessor stoddardi (Emery) ; Wheeler and Creighton, 1934: 385 (first combination in Veromessor ).
Novomessor ( Veromessor) stoddardi (Emery) ; Enzmann, 1947: 152 (revived combination in Novomessor [ Veromessor ], in key).
Veromessor stoddardi (Mayr) ; Creighton, 1950: 157 (revived combination in Veromessor ).
Messor stoddardi (Emery) View in CoL ; Bolton, 1982: 341 (first combination in Messor View in CoL ).
Veromessor stoddardi Ward, Brady, Fisher, and Schultz, 2015: 13 (revived combination in Veromessor ).
Worker diagnosis. This species is uniquely characterized by the following combination of features: (1) head and mesosoma orangish-red to orangish-brown, gaster blackish-orange to blackish, (2) medial lobe of clypeus lacking medial groove but with several lateral longitudinal rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4) antennal scape narrowest immediately distad of basal bend, (5) MOD distinctly less than OMD, OI <25.0, (6) cephalic dorsum moderately shining between fine, widely spaced, longitudinal rugae; antennal fossa lacking concentric rugae, (7) psammophore poorly developed; ventral surface of head capsule with scattered straight or evenly curved hairs, but J-shaped hairs mostly absent, (8) dorsum of pronotum with fine, wavy to weakly irregular, longitudinal rugae; sides of pronotum with longitudinal rugae that fade to disappear posterad, posterior one-third often granulate; mesonotum mostly smooth and shining, lacking rugae or with longitudinal rugae that are distinctly weaker than those on dorsum of pronotum; mesopleura with rugae that traverse longitudinally to posterodorsally, interrugae weakly coriarious to moderately granulate, (9) propodeal spines short, acuminate, length less than distance between their bases; infraspinal facet and propodeal declivity rugose or weakly to strongly coriarious, weakly to moderately shining, and (10) metasternal process large, narrowed laterally, slightly higher than long with angulate apex; anterior and posterior surfaces decline steeply; partly translucent ( Figures 48–49 View FIGURE 48 View FIGURE 49 ).
Measurements. lectotype ( n = 66). HL 1.72 (1.03–1.80); HW 1.80 (0.93–1.83); MOD 0.33 (0.21–0.33); OMD 0.44 (0.23–0.44); SL 1.22 (0.83–1.45); PNW 1.02 (0.62–1.11); HFL 1.63 (0.92–1.71); ML 2.20 (1.27–2.20); PW 0.34 (0.19–0.38); PPW 0.50 (0.26–0.52). Indices: SI 67.68 (66.85–100.69); CI 104.65 (87.07–109.20); OI 18.33 (16.09–24.75); HFI 90.56 (68.03–114.85).
Queen diagnosis. This caste is diagnosed by the following combination of features: (1) head dark orangishbrown to reddish-brown, gaster slightly lighter; pronotum, dorsal portion of anepisternum, mesoscutellum, propodeum dark orangish-brown; rest of mesosoma dark blackish-orange to blackish, (2) medial lobe of clypeus lacking medial groove but with several lateral longitudinal rugae, (3) mandibles with 8 teeth, (4) dorsal surface of base of scape slightly flattened; maximum basal width of scape similar to maximum preapical width, (5) MOD slightly greater than OMD, (6) cephalic dorsum with fine, wavy to irregular longitudinal rugae that fade to disappear near posterior margin; interrugae moderately shining, (7) psammophore poorly developed, (8) sides of pronotum weakly to moderately punctate between fine longitudinal rugae; mesoscutum and mesoscutellum smooth and shining with scattered piligerous punctures; anepisternum shining, weakly punctate between fine longitudinal rugae, both rugae and punctuation weak to absent ventrad; katepisternum mostly shining and superficially roughened with short, longitudinal striae anterad and posterad, (9) sides of propodeum weakly shining and densely punctate between longitudinal and oblique rugae; propodeal spines triangular, acuminate, distinctly shorter than distance between their bases; infraspinal facet shining, superficially punctate above, propodeal declivity smooth and shining, and (10) metasternal process large, narrowed laterally, slightly higher than long with angulate apex; anterior and posterior surfaces decline steeply; partly translucent ( Figure 50 View FIGURE 50 ).
Measurements. ( n = 6). HL 1.56–1.72; HW 1.61–1.75; MOD 0.40–0.44; OMD 0.34–0.41; SL 1.20–1.32; HFL 1.61–1.69; ML 2.77–2.97; PW 0.39–0.45; PPW 0.61–0.67. Indices: SI 74.07–76.97; CI 96.99–110.76; OI 24.12–26.71; HFI 93.71–101.24.
Male diagnosis. This caste is diagnosed by the following combination of features: (11) blackish brown, appendages light brown, (12) medial lobe of clypeus abruptly descendant distad, disc weakly shining with coarse oblique to longitudinal rugae, (13) mandibles with 3–4 teeth basad of preapical tooth, (14) anterior ocellus above level of top of eyes, (15) anepisternum weakly shining, finely punctate between closely spaced, moderately coarse, longitudinal rugae; katepisternum shinier, disc weakly roughened, posterad with moderately coarse oblique to longitudinal rugae, (16) propodeum densely punctate between coarse, mostly longitudinal rugae, interrugae weakly shining; propodeal spines triangular, acuminate, length less than distance between their bases, (17) metasternal process distinctly higher than long, apex narrowly rounded, and (18) subpetiolar process short, elongate-triangular to spiniform or digitiform, apex acuminate to rounded ( Figures1J View FIGURE 1 , 51 View FIGURE 51 ).
Measurements. ( n = 2). HL 1.15–1.19; HW 1.12–1.15; MOD 0.45–0.46; OMD 0.11–0.14; SL 0.46–0.52; HFL 1.96–2.07; ML 3.07–3.12; PW 0.46–0.49; PPW 0.83–0.90; AOD 0.12–0.13; IOD 0.32–0.32; OOD 0.40–0.42. Indices: SI 41.07–45.22; CI 94.12–100.00; OI 40.00–40.18; HFI 175.00–180.00.
Additional material examined. MEXICO: Baja California: 38.5 mi NW Catavina at 1 mi N El Progreso , Feb 10, 1993 ( RAJC; UCDC) ; Sierra San Borja at 9.9 mi N San Borja, 1970’, Mar 11, 1998 ( RAJC) ; 4.4 mi W Meling Ranch , 2450’, Mar 25, 2001 ( CIDA; NHMW; RAJC; UCDC) ; Meling Ranch , 2200’, no date (not examined; R. Snelling, pers. obs.) ; 4 mi S Misión Descanso , 700’, May 14, 1952 ( LACM; UAIC) ; Rosarito Beach , Jun 14, 1938 ( CASC; LACM; USNM) ; 19 mi N Ensenada , 1100’, May 15, 1952 ( LACM; USNM) ; 4 mi S La Misión , 700’, May 14, 1952 ( USNM) ; Rancho Cañadas at 13 km NE Camalú, 260 m, Aug 26, 1994 (not examined; M. Bennett, pers. comm.) ; 2.8 mi E San Telmo , Jul 14, 1979 ( UCDC) ; 14 km ENE Jct Hwy 1 at Road to Bahía de los Ángeles, 480 m, Apr 7, 1998 ( UCDC) ; “ The Oaks ” at 5 mi N Santo Thomas, May 9, 1999 ( UCDC) ; 10 mi E San Quintin , no date (not examined; R. Snelling, pers. obs.) . UNITED STATES: California: Monterey Co.: FS Hastings Natural History Reserve, Apr 25, 1941 ( LACM) ; Paraiso Springs , no date (not examined; R. Snelling, pers. obs.); no loc, Jun 1963 ( USNM) . Riverside Co.: Cleveland National Forest, Forest Service Rd 8S01 at 3 mi E Jct 8S01 & 8S04 (W of Lake Elsinore ), May 7, 1984 ( LACM) ; Lake Skinner , 462 m, Jul 29, 1996 ( UCDC) . San Bernardino Co.: Chino Hills State Park at 6 km S Los Serranos, 250 m, Jun 13, 1994 (not examined, M. Bennett, pers. comm.) . San Diego Co.: San Isidro, 200’, May 13–15, 1952 & Jul 9, 1952 ( LACM; USNM) ; Descanso , Feb 1898 ( USNM) ; 5 mi E Descanso, 3000’ (not examined, R. Snelling, pers. obs.); Santa Ysabel Ecological Reserve , 957 m, Jun 2003 ( UCDC) ; Sweetwater River , 100 m, May 29–Jun 3, 2003 ( UCDC) ; Sycamore Crossing, Peñasquitos Canyon , 25 m, May 1, 2004 ( UCDC) . Santa Barbara Co.: 4 km E San Marcos Pass , 980 m, May 5, 1986 ( JTLC) ; 21 km ESE Santa Ynez, 980 m, May 5, 1986 (not examined, M. Bennett, pers. comm.) ( Figure 52 View FIGURE 52 ).
Etymology. The specific epithet, stoddardi , was presumably derived from someone named Stoddard, but no reference to this person has been located.
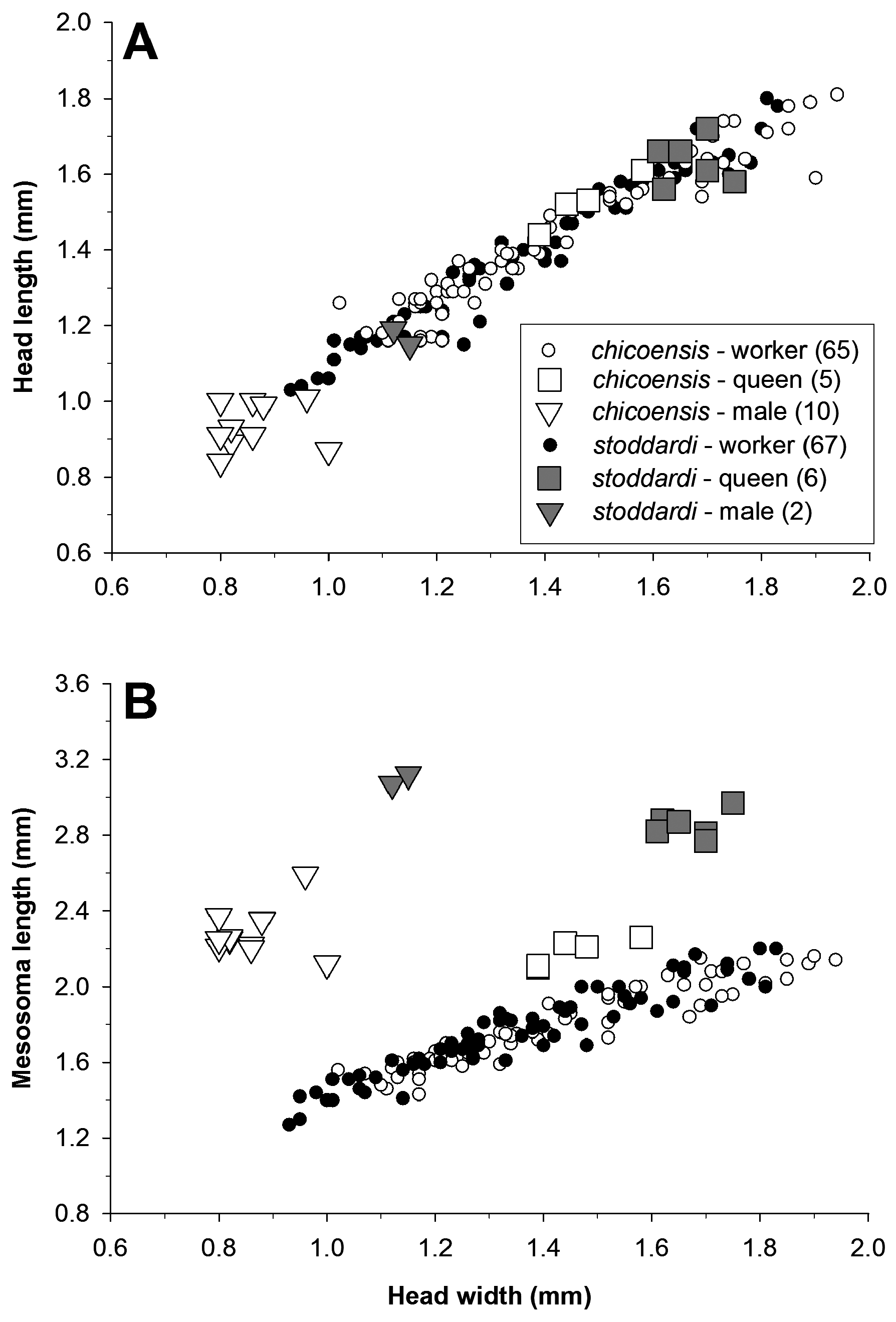
Discussion. The poorly developed psammophore distinguishes V. stoddardi from all congeners except V. andrei and V. chicoensis . Veromessor andrei is the only congener with a poorly developed psammophore that is likely to cooccur with V. stoddardi . The two species can be distinguished based on: (1) V. stoddardi has short propodeal spines (length less than the distance between their bases), (2) strongly polymorphic workers, and (3) weak, regular rugae on cephalic dorsum posterior to eyes and dorsum of pronotum. In V. andrei : (1) propodeal spines long (length> 3× the distance between their bases), (2) workers mostly monomorphic, and (3) notably coarse rugae on cephalic dorsum posterior to eyes and dorsum of pronotum. Veromessor stoddardi is most similar to the closely related species V. chicoensis , from which it can be distinguished by: (1) antennal scape narrowest immediately distad of basal bend; maximum basal width of scape about equal to maximum preapical width, and (2) in dorsal view, the mesonotum usually smooth and shining, rugae distinctly weaker than those on pronotum. In V. chicoensis : (1) antennal scape narrowest near midlength; maximum basal width of scape greater than maximum preapical width, and (2) in dorsal view, rugae on mesonotum similar to those on pronotum, interrugae weakly dull (also see discussion under P. chicoensis ).
Workers of these two species display few consistent morphological differences, but queens and males differ in both morphology and size. Worker allometry is similar for both species (see above), but queens and males of V. stoddardi are distinctly larger than those of V. chicoensis ( Figure 19 View FIGURE 19 ). A molecular phylogeny based on UCEs shows that both species form divergent, reciprocally monophyletic lineages (M.L. Borowiec, unpub. data).
Wheeler and Creighton (1934) wrongly restricted the type locality of V. stoddardi to San Diego County, indicating, “the type series comes from several localities all in San Diego County, California. It seems preferable, for this reason to regard San Diego County as the type locality.” This is a remarkable statement given that none of the type material came from San Diego County. The locality cited in the description by Emery (1895) is San Jacinto, Riverside County, California, and this is the correct type locality, as noted by Creighton (1950).
Biology. Workers of V. stoddardi are solitary foragers that commence activities near dusk and forage through the night (M. Bennett, pers. comm.). Another observer indicated that colonies have long foraging columns (K.L. Jorda, pers. comm.), such that more information is needed to understand foraging biology of this species. Colonies probably contain 1,000 –2,000 workers ( Creighton, 1953; R.A. Johnson, pers. obs.). Workers are strongly polymorphic. Nests are often placed in hard clay soil and often have two to three entrances ( Creighton, 1953).
Gland chemistry has not been examined in V. stoddardi . Like other small-colony congeners, workers of V. stoddardi have a small pygidial gland reservoir and lack a textured tergal cuticle ( Hölldobler et al., 2013).
Mating flights occur near dusk and post-dawn hours in June and July ( Creighton, 1953; M. Bennett, pers. comm.). Nothing is known about mating frequency or colony founding.
Veromessor stoddardi occurs at elevations from 60–980 m, and it appears to be mostly restricted to soils with higher clay content (M. Bennett, pers. comm.). This species occurs in the Baja California desert, California coastal sage and chaparral, and California montane chaparral and woodlands ecoregions, as defined by Olson et al. (2001) ( Figure 52 View FIGURE 52 ).
Additional notes
We also note a putative new species of Veromessor that is unavailable to describe until additional material is collected. Two workers of this species were collected from the northern end of the Mohawk Dunes, Yuma County , Arizona, during summer (year unknown). These specimens were examined by Roy Snelling, who stated that they were a new species closely related to V. pergandei . These two specimens were undoubtedly deposited at LACM, but the senior author could not find these workers during a visit to LACM in 2018, and they are presumed lost. The senior author and others (including RRS) have revisited Mohawk Dunes multiple times through the years without finding additional specimens, and we write this addendum so that other collectors can be on the lookout for this species. We presume that this is a small-colony, nocturnal species given the difficulty finding additional specimens. As a note on geographic range, there are numerous sand dunes in the area, and we presume this species would also occur south into the sand dunes of the Gran Desierto Altar in northwestern Sonora, Mexico. We predict ecology of this species ( Table 1 View TABLE 1 and below), which we list as V. sp. cf. pergandei .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Myrmicinae |
|
Genus |
Veromessor stoddardi
| Johnson, Robert A., Borowiec, Marek L., Snelling, Roy R. & Cole, Arthur C. 2022 |
Stenamma ( Messor ) stoddardi
| Emery, C. 1895: 307 |