
Listriodon retamaensis, Pickford & Morales, 2003
|
publication ID |
https://doi.org/10.5281/zenodo.5377612 |
|
persistent identifier |
https://treatment.plazi.org/id/038987C1-F355-FFD2-9914-FA24FE4FCCF9 |
|
treatment provided by |
Marcus |
|
scientific name |
Listriodon retamaensis |
| status |
sp. nov. |
Listriodon retamaensis n. sp.
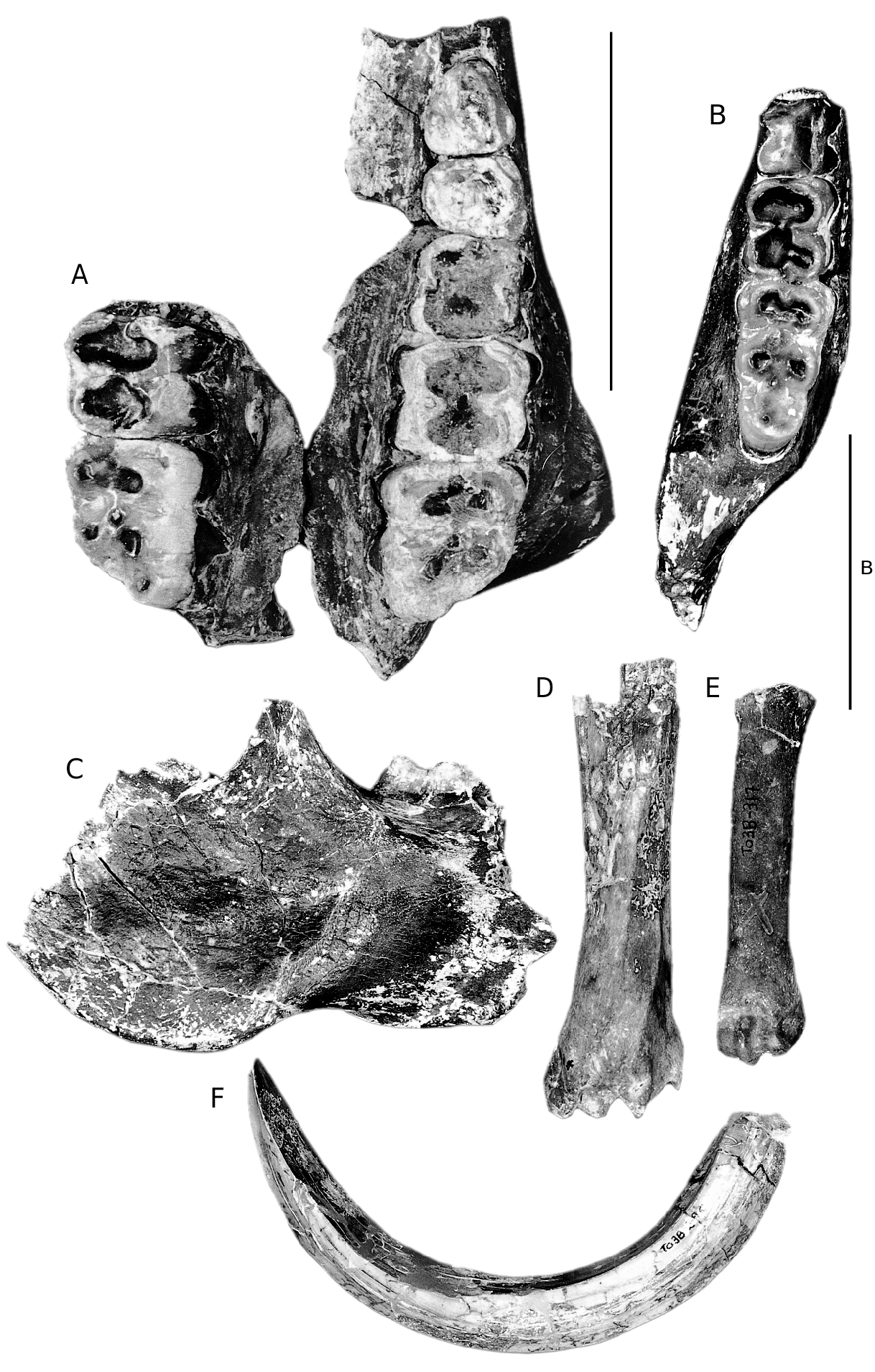
HOLOTYPE. — RET 574 ( Fig. 1G View FIG ), snout of a female individual with left I3, left and right canine roots, left and right P1-P3 (left P3 broken), housed at the Museo Nacional de Ciencias Naturales , Madrid.
ETYMOLOGY. — Named for the type locality.
PARATYPES. — RET 740 , right I1 ( Fig. 1A View FIG ) ; RET 348 , left maxilla with P3-M3 and right maxilla with M2- M3 ( Fig. 2A View FIG ) ; RET 832 , upper male canine ( Fig. 1B View FIG ) ; right p4 ( Fig. 1C View FIG ); RET 2 , left mandible fragment with m1-m3 (occludes perfectly with RET 348 ) ( Fig. 2B View FIG ) ; RET 338 , right mandible fragment with broken m3 ( Fig. 2C View FIG ). Housed at the Museo Nacional de Ciencias Naturales , Madrid .
TYPE LOCALITY. — Retama, Huete, Spain ( MN 4a).
OTHER LOCALITIES. — Spain: Acacias ( MN 4b); Casal das Chitas ( MN 4b); Corcoles ( MN 4a); Estacion Imperial ( MN 4b); La Hidroelectrica ( MN 4b); Tejar Manzanares ( MN 4b or MN 5); Munebrega ( MN 4b); Olival da Susana ( MN 4b); Quinta Grande ( MN 4a); Quinta das Flamengas ( MN 4b); Quinta de Lobeira ( MN 4a); Quinta da Raposa ( MN 4a); Quinta da Silvéria ( MN 4b); Terrero ( MN 4a); Torralba ( MN 4b); Valdemoros ( MN 4b). France: Bézian ( MN 4b).
AGE. — This new species of Listriodon is known from several middle Miocene localities in Europe ranging in age from MN4a to MN5 ( Figs 3 View FIG ; 4 View FIG ) (c. 17-15 Ma).
DIAGNOSIS. — A small species of Listriodon with moderately elongated snout (diastemata between canine and premolar row shorter than in Eurolistriodon ), full complement of upper premolars, three lobed I1 with a length-width index of c. 177; bunolophodont dentition; metapodials short and robust.
REMARKS
Van der Made (1996) listed material from Echzell and?Georgensmund as Bunolistriodon aff. latidens ( Biedermann, 1873) , but the material falls within the range of variation of L. latidens and L. lockharti Pomel, 1848 and it is more likely to belong to one or other of these taxa rather than to L. retamaensis n. sp. Some specimens from MN4a were also assigned to Bunolistriodon aff. latidens by Van der Made (1996), but these possess a single lobed upper central incisor, and are here assigned to Eurolistriodon adelli . Thus, apart from one locality in France (Bézian) all the known occurrences of L. retamaensis n. sp. are from the Iberian Peninsula.
DESCRIPTION
Skull
In palatal view ( Fig. 1G View FIG ) the anterior end of the snout is oval in outline, being widest at the canines. There is a distinct waist behind the P1, beyond which the palate widens gradually to where it is broken behind the P3. The incisive foramina are located immediately behind the I1 and extend backwards as far as the distal margin of the alveolus of I3. The maxillo-palatine sutures run almost parallel to each other distally from the rear of the incisive foramina. On each side of the palate a groove runs backwards from the incisive foramen coursing close to the alveolar processes. The suture between the premaxillae and maxillae is raised into a slight but sharp ridge.
The incisor alveoli form a curved battery, the I1 being the largest and the I3 the smallest of the incisors. A short diastema separates the I3 from the canine, the margin of the palate being curved dorsally, thereby forming a shallow niche for the lower canine. The P1 is a two-rooted tooth separated from the rear of the canine by a very modest gap. Behind it there is a short diastema marked laterally by a depression in the outer surface of the maxilla. The P2 and P3 are in contact with each other.
In lateral view, the anterior margin of the premaxilla sweeps upwards steeply from the alveoli of the central incisors, the hindmost margin of the anterior nares being above the alveolus of I3. The anterior ends of the nasal bones are broken but probably did not extend very far anteriorly.
Inside the nasal cavity of the Retama snout, above and behind the level of the P2 there is a well developed sheet of bone oriented vertically which separates the nasal cavity into two halves. In distal view, a curved sheet of bone is seen to extend into the nasal cavity from about half the height of the snout while a second sheet curves into the cavity from the roof of the snout near the suture between the maxilla and the nasals. In a remnant of limestone that is still lodged in the nasal cavity it is possible to discern portions of thin curved sheets of bone that represent turbinates. These extend as far forwards as the level of the canine alveoli, if not further forwards, from which it is deduced that the sense of smell was well developed in this species.
RET 348 ( Fig. 2A View FIG ) is a fragmented left maxilla containing P3-M3 and part of the right maxilla with M2-M3. The midline of the maxilla is not preserved, nor is its distal margin. It is not possible to determine the width of the palate, nor its extension behind the molar row. The palatal foramen is located close to the disto-lingual root of the M2. Close behind the M3 there is a well developed tubercle of bone above which is a groove which probably housed a blood vessel which coursed from the palate upwards and outwards behind the M3.
Upper dentition ( Table 1)
The upper central incisor, RET 740 ( Fig. 1A View FIG ), is slightly worn on its lingual aspect, but is otherwise well preserved. The tooth is wide mesiodistally and its apical edge is divided into three lobes, the central one being the narrowest. Its length-width index is 177 (for calculation of index, see Van der Made 1996). There is a prominent lingual cingulum which runs the entire length of the tooth. The root is appreciably narrower than the crown and it tapers rapidly towards its apex which has been broken away.
The I 3 in situ in the snout is a small tooth with a triangular crown in lingual view. The anterior edge is worn flat parallel to the palate while the distal edge is unworn. It has a weakly developed, but sharp, lingual cingulum.
The canine roots are oval and short, indicating that this individual was probably a female. The P1 is a two-rooted tooth, the roots splaying out rapidly so that the length of the premolar alveoli is greater than that of the crown. The crown is bunoid with a central cusplet from which low crests run anteriorly and distally. Lingually there is a low, rounded cingulum. The P2 is more than twice the size of the P1 and its crown is formed of a main centrally positioned cusp behind which is a low shelf and a small distal cusplet. Ridges run anteriorly and distally from the apex of the main cusp. A prominent wear facet caused by abrasion with the lower second premolar scores the anterior crest of the P2. In occlusal view the P3 is a large triangular tooth. The main cusp and its anterior and distal crests are obliquely oriented and the disto-lingual corner of the tooth is formed of a large cusplet which is separated from the main cusp by a valley. Labially, the distal part of the crown is adorned by a cingular ridge. The anterior surface of the tooth is the site of a large flat wear facet caused by abrasion with the p3.
An unworn upper male canine ( Fig. 1B View FIG ) has an almost circular cross section with low pre- and post-crista. The enamel is scored by slightly wavy, shallow, longitudinal grooves. The pulp cavity is visible where part of the root has broken away, revealing that this tooth was not as elongated nor as curved as the upper male canines that typify Eurolistriodon , and later species of Listriodon .
In the maxilla RET 348, the P4 is moderately worn but it is possible to make out that the buccal cusp was probably formed of a pair of closely appressed cusps. Lingually there is a single cusp bordered by a well developed cingulum which extends along the anterior, lingual and distal margins of the crown and reaches onto the labial surface of the tooth anteriorly and distally. The upper first and second molars in this specimen are so worn that little morphology is preserved. It is possible to determine, however, that both teeth possessed prominent buccal cingula. The M3 possesses four main cusps and a lingually positioned talon. It has well developed but low anterior and median accessory cusplets. The four main cusps and the two accessory cusplets all have dentine exposed at their tips. The dentine exposures of the anterior cusps are joined to that of the anterior accessory cusplet. The buccal margin of the crown possesses a well developed cingulum. The tooth, even though well worn, is evidently of the bunolophodont type.
Mandible
RET 338 and RET 2 ( Fig. 2B, C View FIG ) are parts of the right and left bodies of an individual lower jaw which occlude perfectly with the two maxillae, RET 348. RET 2 is so fragmentary that it reveals little about mandibular morphology except that the root of the ascending ramus sweeps upwards behind the m3 as in other listridonts, and in contrast to the short mandibles of hyotheres. RET 338 in contrast preserves much of the body below the m3 including the angle and part of the
Upper dentition Length Breadth AR 278 left P3 15.1 -
AR 258 left P3 17.0 13.9 ART 56 right P3 16.0 14.5 ART 56 right P4 13.0 15.6 AR 275 left P4 13.0 15.2 AR 289 left P4 13.0 16.0 AR 277 right M1 18.7 16.4 AR 276 right M1 18.1 17.8 AR 289 left M1 18.7 17.2 AR 118 left M1 18.7 16.8 AR 289 left M2 21.0 20.9 AR 289 left M3 25.5 22.0 ART 59 right M3 21.5 19.2
masseteric fossa. The lower border of the jaw below the third molar terminates in a prominent flange and lingual tubercle which is separated from the slightly descending angle by a low crest of bone. The internal and external surfaces of the jaw distal to the third molar are marked by well developed rugosities representing muscle attachments. The lower border of the masseteric fossa is everted to a small extent. The mandibular foramen enters the lingual side of the ascending ramus 30 mm behind the talonid of m3.
Lower dentition ( Table 1)
The p4 ( Fig. 1C View FIG ) is rectangular in occlusal outline. Centrally there is a prominent main cusp beside which is a large, offset innenhugel. Anteriorly the anterior crest of the main cusp is swollen and descends rapidly towards a well developed but low anterior cingulum. Distally there is a prominent talonid cusp joined lingually and labially by a swollen cingulum.
The lower first and second molars of RET 2 are deeply worn and the most informative morphological characters have been eradicated. The m3 is also deeply worn but it is possible to make out that it consisted of five main cusps with median and posterior accessory cusplets. The main cusps are arranged in two loph-like pairs with a centrally positioned talonid distally, and there is an anterior cingulum. In lateral view the cervix of the tooth is seen to ascend distally. The five main cusps and the two accessory cusplets all have dentine exposed at their tips, that of the anterior cusp pair forming a loph-like dentine lake. In RET 338 only the talonid and the broken surface of the distal cusp pair of the m3 are preserved.
The dimensions of the Retama teeth fall outside the currently documented range of metric variation of Listriodon lockharti ( Pomel 1848) and Listriodon latidens ( Biedermann, 1873) ( Van der Made 1996; Van der Made et al. 1998), but they are close in size to the teeth of Eurolistriodon adelli as noted by Van der Made (1996) (Figs 5- 10). However, morphologically they fall well outside the known range of variation of the latter species, their cheek teeth being more lophodont and the upper central incisor possesses three lobes rather than one. It is impossible to determine from metric data alone to which species isolated cheek teeth of small listriodonts may belong, the size distribution of all species ( adelli , retamaensis , lockharti and latidens ) overlapping each other. For this reason, we consider that some of the material previously assigned to L. aff. latidens ( Van der Made 1996) belongs to E. adelli while some of it belongs to small individuals of L. lockharti . The main differences between the cheek teeth of these listriodonts concern the degree of bunodonty and lophodonty. For example, the molars of E. adelli are more bunodont than those assigned to L. lockharti , but the distinctions can disappear with moderate wear. The upper central incisors of the two groups are different, as is the morphology of the canine flange and there are important differences in the postcranial skeleton ( Pickford & Moya Sola 1995). From the morphological data we conclude that there are two genera of listriodonts in Spanish deposits ranging in age from MN4a to MN5.
COMPARISONS AND DISCUSSION
The Retama snout closely resembles a specimen from Bézian attributed to Bunolistriodon lockharti by Ginsburg & Bulot (1987) but identified as Bunolistriodon aff. latidens by Van der Made (1996). It differs markedly from the species Eurolistriodon adelli from Els Cassots, Spain ( Pickford & Moya Sola 1995) which is a long snouted listriodont in which the anterior premolars have been suppressed, in which there is a long post-canine diastema and in which the molars are
Pickford M. & Morales J.
more bunodont. It is unlikely that these differences are due solely to sexual dimorphism within a single species, even though in listriodonts sexual dimorphism of the snout region is marked ( Pickford & Moya Sola 1995).
At the time of their study, Van der Made (1996, 1997a) and Van der Made & Alferez (1988) recognised that localities such as Corcoles and several other Iberian sites of similar age (MN4) possessed two listriodont species, but the avail-
European
Age
Land mammal
Ma France Germany Switzerland Zones Paratethys Mideast Indian China Afro-Arabia & Med. Subcon.
able material was not well enough preserved to permit the formal naming of a new taxon. In a note printed at the end of his monograph, Van der Made (1996) suggested that the material assigned by him to B. aff. latidens belongs to Bunolistriodon adelli a species which was published while his monograph was in preparation. The snout from Retama differs markedly from that of E. adelli , and reveals that by MN 4 there were in fact three distinct listriodonts in Spain: Eurolistriodon adelli , Listriodon retamaensis n. sp. and Listriodon lockharti .
It now appears that by the onset of MN 4, there were two genera of listriodont suids in Europe, one which possessed a derived pattern of bunolophodont cheek teeth, trilobed upper central incisors and a canine which emerged from the end of the canine flange, represented by two species ( L. lockharti and L. retamaensis n. sp.), and one which retained a plesiomorphic pattern of bunodont (crystodont) cheek teeth, a single lobed upper central incisor, and upper canines which emerged from the ventral surface of the canine flanges ( Eurolistriodon adelli ). The two genera are further distinguished by their postcranial skeletons – short and robust metapodials in Listriodon retamaensis n. sp. and L. lockharti , compared with long and gracile ones in Eurolistriodon adelli . The two genera share several derived features of the skull and dentition including
Pickford M. & Morales J. FIG. 5. — Size variation (in mm) of the upper dentition of Listriodon retamaensis n. sp. Symbols:, maximum and minimum measures, various localities;l, range of variation.
distal prolongation of the palatines, elongated snout with diastemata between the canines and premolars, spatulate anterior palate, and mesiodistally elongated upper central incisors, indicating that they diverged from a common stock some time prior to MN 4.
The morphology and length/breadth index of the upper central incisors of L. retamaensis n. sp. are close to those of L. lockharti , and differ from those of E. adelli and L. latidens , as well as from those of other Listriodon species. The Retama cheek teeth are more lophodont than those of E. adelli , and resemble those of both L. latidens and L. lockharti . It would thus appear that the listriodonts from Retama were drawn from an early population of Listriodon , the body size of which was smaller than later populations of the genus.
Ginsburg & Bulot (1987) described a snout from Bézian, France, which they identified as Bunolistriodon lockharti in which the nasals are complete, extending forwards only as far as the level of the I3 alveolus. The canine jugum is modestly developed, there being no sign of a canine flange, but this may well be because the individual is female. In section, the snout is rounded dorsally with a flat palatal floor. This specimen is here assigned to Listriodon retamaensis n. sp.
Sexual dimorphism
It could be argued that the morphological differences between Eurolistriodon adelli and Listriodon retamaensis n. sp. merely represent sexual differences, the material assigned to E. adelli being male and that assigned to L. retamaensis n. sp. being female. However, the differences between the two forms are considerably greater than is usually accepted as being due to sexual dimorphism in suids, and they comprise cranio-dental and postcranial parts of the skeleton. It would,
Length i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 Breadth
FIG. 6. — Size variation (in mm) of the lower dentition of Listriodon retamaensis n. sp. Symbols:, maximum and minimum measures, various localities;l, range of variation.
for instance, be extremely doubtful that males would suppress the anterior premolars while females retained them or that males would possess more bunodont cheek teeth than females, or that the upper central incisors of males would have one lobe while those of females had three. The post-canine diastema in Eurolistriodon is long (65 mm in the holotype) while that in L. retamaensis n. sp. is shorter. Even if we take the homologous measure of the distance between the rear of the canine and the front of P3, it is evident that the Retama snout, with a distance of 38 mm, is considerably shorter than that of Eurolistriodon adelli which has a gap of 65 mm. The Inönü, Turkey ( MN 6), male palate assigned to L. latidens ( Pickford & Erturk 1979) has a C-P3 measure of about 38 mm, appreciably shorter than that of E. adelli . Whilst it is admitted that the length of diastemata can vary appreciably within suid populations ( Van der Made 1991, 1997b) it seems excessive to consider such large differences to be due solely to individual variation, especially when they are considered in conjunction with other differences in morphology. If we now move on to take into account the morphology of the canine flange, we notice even more striking differences between Eurolistriodon and Listriodon . In E. adelli the canine flange in the male is extremely wide – it doubles the breadth of the maxilla – and the canine emerges from the ventral aspect of the flange, not at its tip. In all Listriodon specimens where this part of the anatomy is preserved, the canines emerge from the ends of the canine flanges. Added to this is the evidence afforded by the postcranial skeleton, in particular the long and slender metapodials of Eurolistriodon adelli which contrast with the shorter more robust metapodials of Listriodon retamaensis n. sp. and other species of Listriodon . For all these reasons, we consider it likely that
Pickford M. & Morales J. FIG. 7. — Size variation (in mm) of the upper dentition of Listriodon lockharti Pomel, 1848 . Symbols:, maximum and minimum measures; l, range of variation;´, Retama fossils.
there are two distinct genera of listriodont suids ( Eurolistriodon and Listriodon ) in the basal middle Miocene deposits of Europe.

| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |