
Danionella cerebrum, Britz & Conway & Rüber, 2021
|
publication ID |
https://doi.org/ 10.1038/s41598-021-97600-0 |
|
publication LSID |
lsid:zoobank.org:pub:65198876-420A-4C63-B953-F84C55F3F1FB |
|
DOI |
https://doi.org/10.5281/zenodo.5524642 |
|
persistent identifier |
https://treatment.plazi.org/id/EB6CE17B-2A2E-4773-82A4-8EB7A2D40C45 |
|
taxon LSID |
lsid:zoobank.org:act:EB6CE17B-2A2E-4773-82A4-8EB7A2D40C45 |
|
treatment provided by |
Carolina |
|
scientific name |
Danionella cerebrum |
| status |
sp. nov. |
Danionella cerebrum new species.
Holotype. BMNH 2021.8.30.1 , female, 12.6 mmSL, Myanmar, Yangon Division, Hmawbi , roadside canal draining into Tandabin Chaung, 17° 06.200′ N 96° 02.890′ E, Britz et al., 18 Oct 2008. GoogleMaps
Paratypes. MTD 39985 , 245 specimens, 7.5–12.0 mm SL, same information as holotype. GoogleMaps MTD 39986, 20 c&s, 10.5–13.5 mm SL, same information as holotype. GoogleMaps ZRC 62210, 36, 7.2–12.4 mm SL, same information as holotype. GoogleMaps NRM 71156, 36, 7.7–10.8 mm SL, same information as holotype. GoogleMaps USNM 439009 , 36, 7.8–12.0 mm SL, same information as holotype. GoogleMaps BMNH 20.21.6.2.1-200 , 200, 7.2–11.8 mm SL, same information as holotype. GoogleMaps MTD 39987, 1, 10.4 mm SL, c&s, Myanmar, Bago Division, Daik U, Daikme Chaung , 17° 48.267′ N 96° 39.826′ E, Britz et al., 19 Oct 2008. GoogleMaps MTD 39988, 10– 12 mm SL, 4, unnamed stream 14.4 km NE of Daik U, 17.89314° N 96.56361°E, Britz et al., 19 Oct 2008. GoogleMaps MTD 39989, 1 c&s, 11.4 mm SL, unnamed stream 14.4 km NE of Daik U, 17.89314° N 96.56361°E, Britz et al., 19 Oct 2008. GoogleMaps
Additional material (non type): MTD 39990 , 29 specimens, 7.8–11.5 mm SL, aquarium shop in Bago, reportedly from Tandabin Chaung, Britz et al., 19 Oct 2008. MTD 39991, 8 c&s, 8.5–11.3 mm SL, aquarium shop in Bago, reportedly from Tandabin Chaung, Britz et al., 19 Oct 2008.
Diagnosis. Danionellacerebrum isdistinguishedfrom D. translucida , D. dracula , and D. priapus bythenumber of anal-fin rays (15–18 vs. 12–15 in D. translucida , 12–14 in D. dracula , 20–21 in D. priapus , Table 1 View Table 1 ). It is further distinguished from D. mirifica , D. dracula , and D. priapus by fewer vertebrae (33–35 vs. 36–38, Table 1 View Table 1 ), from D. priapus and D. dracula by fewer pectoral-fin rays (6 vs. 8 in D. priapus and 7 in D. dracula , Table 1 View Table 1 ), from D. translucida and D. dracula by the presence of a ventromedially directed cartilage flange from the taenia marginalis anterior that approaches the trabecula communis (vs. absence, Fig. 2c,d), and from D. dracula by the presence in the male of bony flanges on the outer arm of the os suspensorium and a connection of these to the lateral process of vertebra 2 (vs. absence of flanges and of connection to second lateral process), the presence of a maxillo-mandibular cartilage (vs. absence), the absence of odontoid processes in the male (vs. presence), more anal-finpterygiophores (14–17 vs. 11–13, Table 1 View Table 1 ), more principalcaudal fin rays (9 + 9 vs. 8+ 8, Table 1 View Table 1 ) and fewer pelvic-fin rays (5 vs. 6, Table 1 View Table 1 ). Danionella cerebrum can be further distinguished from the similar syntopically living D. translucida , by the last dorsal-fin ray inserted opposite to the last anal-fin ray (vs. last dorsal-fin ray inserted posterior to last anal-fin ray, Fig. 2a,b), by the last anal-fin pterygiophore inserted in front of haemal spine of vertebra 22–24 (vs. 19–21), by the lateral process of the second vertebra blade-like (vs. axe shaped, Fig. 2g,h), and by the distal tip of the fused inner arms of the ossa suspensoria bifurcated (vs. single, Fig. 2e,f) and not reaching the middle of the anterior swimbladder (vs. curving around and reaching middle of anterior swimbladder, Fig. 2e,f).
Description. Maximumknownsize 13.5 mmSL. GeneralbodyshapeillustratedinFig. 1a–d View Figure 1 . Morphometric information based on 10 specimens is provided in Table 2 View Table 2 . Head and eye are large; mouth supraterminal. Nostrils well developed. Lateral line canals and pores on head and body absent. Body elongate with a short dorsal fin, situated opposite to posterior half of long anal fin. Tip of dorsal fin situated posterior to a vertical line through tip of anal fin. Caudal fin furcate with remnants of larval-fin fold in front of its dorsal and ventral margins. A remnant of pre-anal larval-fin fold present in adult females, absent in adult males. Anus and genital papilla of mature males located between pelvic fins, at normal position in front of anal fin in females and in immature males between pelvic and anal fins. Awindow (pseudotympanum) present in body musculature at lateral side of anterior swim bladder chamber, rendering its pigmented surface visible. Scales absent.
Figure 2. Comparisonofskeletalcharactersofclearedandstainedfemalesof Danionellacerebrum , and D. ▸ translucida . Whole skeleton of D. cerebrum (a), MTD 39986, paratype, 11.5 mm SL and D. translucida (b), MTD 39992, 13.1 mm SL in lateral view, illustrating differences in relative position of dorsal and anal fins; vertical lines mark base of anteriormost and posteriormost dorsal-fin ray, respectively, in relation to anal fin. Neurocranium of D. cerebrum, MTD 39986, paratype,13.5 mm (c) and D. translucida (d), MTD 39992, 11.2 mm SL, in dorsal view, star marks ventromedial cartilage flange in (c), which is absent in (d). Tip of fused inner arms of ossa suspensoria in D. cerebrum (e) and D. translucida (f), in ventral view; note bifurcated tip in (e) and single tip in (f). Vertebrae of Weberian apparatus in D. cerebrum (g) and D. translucida (h), in dorsal view; note caudally expanded lateral process in (h), margin marked by line of grey dots. Abbreviations: in, intercalarium;
ios, inner arm of os suspensorium; lp, lateral process of second vertebra; oos, outer arm of os suspensorium; r, rib; sc, scaphium; sw, swimbladder; tr, tripus.
Vertebrae totaling 33(7), 34(23), 35(2), abdominal vertebrae 13(15), 14(16) or 15(1); caudal vertebrae 19(4), 20(17) or 21(11). Ribs present on vertebrae 5–11(30) or 5–12(2). Rib on vertebra 5 dimorphic, stout and well ossified in male, feeble and poorly ossified in female. Dorsal-fin rays 8(31) or 9(1), first two fin rays unbranched (32) and last unbranched (25) or branched (7). Dorsal-fin pterygiophores 7(31) or 8(1). First dorsal-fin pterygiophore inserted behind neural spine of vertebra 18(9), 19(19), 20(3) or 21(1), and last in front of neural spine of vertebra 22(1), 23(22), 24(8) or 25(1). Anal-fin rays 15(8), 16(15), 17(7) or 18(1) with firsttwo rays unbranched (32) and last unbranched (26) or branched (6). Number of anal-fin pterygiophores in front of first haemal spine: 0(7), 1(11) or 2(15). Last anal-fin pterygiophore inserted in front of haemal spine of vertebra 22(14), 23(15) or 24(3). Principalcaudal-fin rays 9 + 9(32) plus 5(2), 6(8), 7(16) or 8 (6) dorsaland 5(1), 6(18), 7(12) or 8(1) ventral procurrent rays. Pectoral-fin rays 6(32) and pelvic-fin rays 5(32).
No visible pigmentation in preserved specimens, except a line above anal-fin base in some specimens. In life, body colourless and largely translucent ( Fig. 1a,b View Figure 1 ), except for a number of melanophores and yellowish colouration covering dorsal surface of skull. Melanophore pattern including an irregular scattering on top and sides of head, a row following the posterior margin of shoulder girdle, oblique melanophore rows along neural and haemal arches and spines in deeper layers of body, a horizontal row along insertion of anal-fin muscles starting above vent and extending posteriorly along caudal peduncle to anterior ventral procurrent rays, a row of melanophores at base of anal fin itself and along first anal-fin rays, and melanophores marking end of the hypural plate. Females with eggs, with numerous, large melanophores in lining of abdominal wall.
Te cleared and double stained specimens ( Fig. 2a,b) revealed that, as in other species of the genus Danionella , the skull, hyopalatine arch, gill arches, endoskeletal shoulder girdle and pterygiophores are mostly cartilaginous with only thin perichondral ossifications giving the skeleton an overall larval appearance. Te following bones are absent in D. cerebrum : kinethmoid, preethmoid, vomer, nasal, parietal, intercalar, extrascapular, infraorbitals 2–5, angular, ectopterygoid, metapterygoid, urohyal, hypobranchials 1–3, posttemporal, postcleithrum, mesocoracoid, pectoral radials 3–4, pelvic radials 1–3, all supraneurals behind supraneural 3, epineurals, epipleurals, uroneural 2, and scales. Exceptions to this theme of bone loss and reduction in the skull are the heavily ossified and toothed ceratobranchial 5, which is essential in food processing in conjunction with the well-ossified basioccipital, which carries the masticatory plate that ceratobranchial 5 works against. Te basioccipital along with the equally wellossified exoccipital houses the intracranial part of the Weberian apparatus (sinus impar, capsules for lagena and asteriscus). Especially well-developed are also the Weberian ossicles and the os suspensorium, whose inner arms are fused in the midline with a bifurcated tip. Tere is a strong sexual dimorphism in the os suspensorium with males having the outer and inner arms more massively developed, the inner arms covering the roof of the swimbladder via posterior processes, the inner and outer arms connected via a broad bony flange and having the outer arms connected to the transverse processes of the second vertebra by a bony process. In addition males have a drumming cartilage associated with the fifh rib and swimbladder. Females lack all these modifications. Also the fifh rib is sexually dimorphic, stout and well ossified in the male and with a ventromedially directed process near its base, and feeble and poorly ossified in the female and lacking such a process.
Etymology. Tespeciesname cerebrum, Latinforbrain , anouninapposition, makesreferencetothefactthat this fish with one of the smallest adult brains among vertebrates has become a promising new model species for neurophysiological studies.
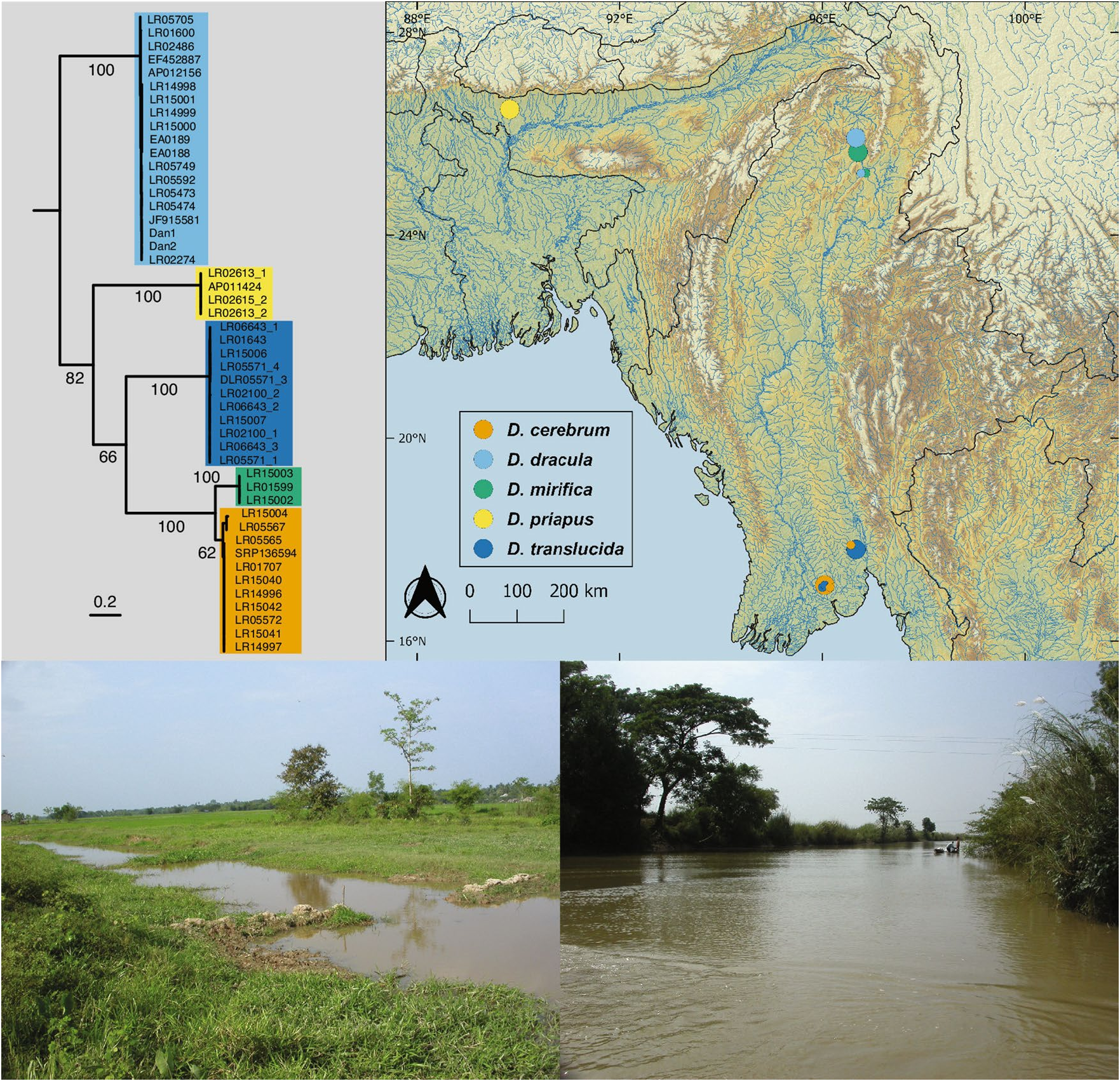
Distribution. Danionellacerebrum isknownfromanumberofstreamsonthesouthernandeasternslopesof the Bago Yoma mountain range ( Fig. 3 View Figure 3 ) of Myanmar: Tandabin Chaung and Bala Chaung in Yangon Division, and from Daikme Chaung (type locality of Danionella translucida ) and an unnamed stream northwest of Daikme Chaung in Bago Division.
Habitat. Allwaterbodiesinwhich Danionellacerebrum wasfound, areturbidlowaltitudestreams ( Fig. 3 View Figure 3 ) with visible flow, surface temperatures of around 30 °C, pH 7.4–7.5 and sof water of 20–100 micro Siemens. Danionellacerebrum was not found at the surface, but at adepth below ca. 30 cm where the water is significantly cooler (ca. 25 °C). Te species was abundant at the type locality in Hmawbi on 18 Oct 2008, where it co-occurred with D. translucida (as evidenced by c&s specimen and DNA sample), which appears to have been less common (a single c&s D. translucida among 28 D. cerebrum ). At Daikme Chaung ( Fig. 3 View Figure 3 ), the type locality of D. translucida , D. cerebrum was uncommon (1 out of 27 c&s specimens of Danionella ) on 19 Aug 2008, but D. translucida was abundant.
Molecularresults. Te 12 individualsof Danionellacerebrum and D. mirifica sequencedforthecytochrome coxidase subunit I (coxI) gene had unique adenin insertions at positions 95 and 624 of the coxI alignment leadingto frame shifs and premature stop codons as previously reported for the gobioid Parapocryptes serperaster 15, the significance of which is unknown. Interestingly, we were also able to identify two adenin insertions in the coxI gene in the whole genome sequence of D. cerebrum provided by Kadobianskyi et al. 16 (GenBank accession SRMA00000000 as D. translucida ) as well as the expected mt DNA tRNAs flanking the coxI, hundreds of base pairs up and downstream of the coxI region sequenced in our study, suggesting a genuine mitochondrial origin of our coxI sequences rather than a nuclear pseudogene (NUMT). For the phylogenetic analyses, however, the two insertions were excluded. In order to make sure that our phylogenetic analyses were not affected by the unlikely inclusion of NUMTs, we also conducted phylogenetic analyses based on an additional mitochondrial marker (16S rRNA) and three nuclear DNA markers (erg2b, rag1, rho). Te different loci resulted in the same tree topology for the five species of Danionella but showed different support values ( Figs. 3 View Figure 3 , 4 View Figure 4 , Electronic Supplementary Fig. 2).
Our initial result based on morphological information that the model organism used in Schulze et al. 11 is not Danionella translucida , but a separate species, D. cerebrum , is supported by our molecular analysis. Samples from three different localities around the southern end of the Bago Yoma mountain range, including the type locality of D. cerebrum , clustered with samples obtained from the stock kept at Bolton Aquarium, from which the individuals usedin Schulze et al. 11, in Penalvaet al. 12 and Kadobiansky et al. 16 originated and with the sample used in Britz et al. 6 and labeled Danionella sp. “South Myanmar ” LR1707.
We found that Danionella cerebrum differs significantly from its close congeners and the uncorrected p-distances in the coxI gene between this species and D. translucida (Electronic Supplementary Table 2 View Table 2 ), the species with which it has been previously confused, ranged from 22.2–22.9%. Even between D. cerebrum and its closest relative D. mirifica , p-distances still range from 10.2–10.9%.
We recovered the following topology among the five species of Danionella independent of the genes analysed ( Fig. 4 View Figure 4 ): ( D. dracula ,( D. priapus ,( D. translucida ,( D. mirifica , D. cerebrum )))). Te age for the split of D. priapus from the remaining three taxa was estimated at ~ 17.9 MYA (95% HPD 14.5–20.9) and the split of D. translucida from D. mirifica + D. cerebrum at ~ 13.3 MYA (95% HPD 10.1–16.1). Te sister taxa D. mirifica and D. cerebrum split at ~ 3.2 MYA (95% HPD 2.0–4.2).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |