
Pogonomyrmex vermiculatus, Emery, 1906
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5033.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4314F784-A510-4F36-9E11-ED1EAC83CEBF |
|
persistent identifier |
https://treatment.plazi.org/id/5027B677-FF1B-A4C9-FF25-FF11FA456F39 |
|
treatment provided by |
Plazi |
|
scientific name |
Pogonomyrmex vermiculatus |
| status |
|
Pogonomyrmex vermiculatus View in CoL
( Figures 3G–H View FIGURE 3 , 106–108 View FIGURE 106 View FIGURE 107 View FIGURE 108 )
Distribution—100C
Pogonomyrmex vermiculatus Emery, 1906: 157 View in CoL (worker). Holotype examined [MSNG: CASENT0280978]. ARGENTINA, Santa Cruz Province: near the Santa Cruz River, 5–7 station (southern Patagonia ). See also Gallardo, 1932: 138, fig. 24.
Pogonomyrmex vermiculatus var. chubutensis Forel, 1913: 218 View in CoL (worker). Holotype [MHNG: CASENT0907120]. ARGEN- TINA, Chubut: mountains of Chubut. See also Gallardo, 1932: 140, fig. 26. NEW SYNONYMY
Pogonomyrmex vermiculatus var. jorgenseni Forel, 1915: 353 (worker). Syntypes examined: 2 workers [MACN], 6 workers [MHNG], 12 workers [MLPA], 1 worker [USNM]. ARGENTINA, Santa Cruz: to the east of Lago Argentina, #284 (Jörgensen leg.). See also Gallardo, 1932: 141, fig. 27; Kusnezov, 1951: 253 (synonymy under vermiculatus var. chubutensis View in CoL ). MACN worker here designated LECTOTYPE [CASENT0281102]. NEW SYNONYMY
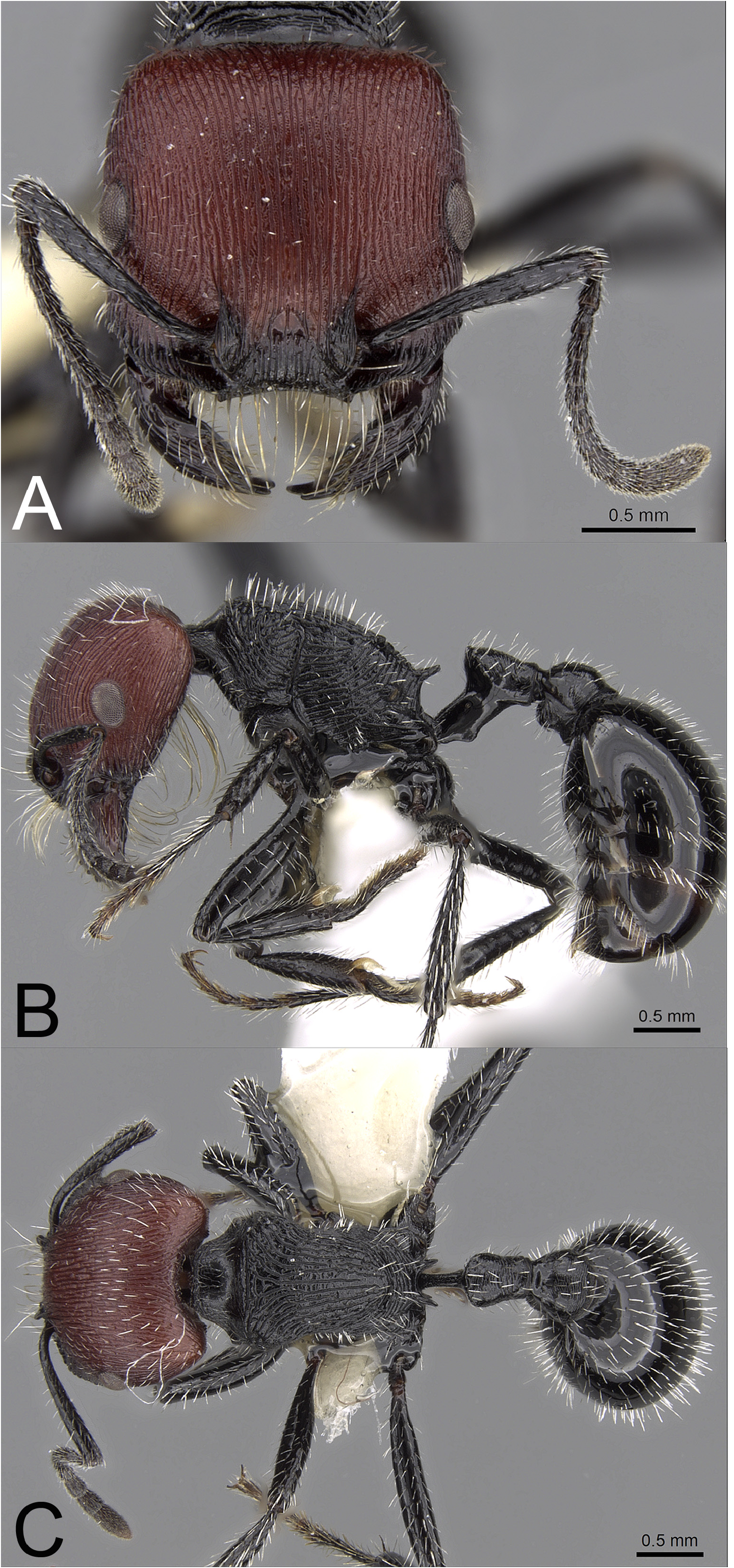
Worker diagnosis. Workers of this species are uniquely characterized by the following combination of features: (1) most workers lack striae on first gastral tergum, rare workers with striae on anterior one-third to one-half of segment, (2) posterior surface of petiolar node and dorsum of postpetiole with transverse rugae, (3) interrugae on cephalic dorsum moderately to strongly granulate, dull, (4) bicolored with head dark reddish-brown to reddish-black, rest of body blackish to black, (5) superior propodeal spines long, length> 0.6–0.8× the distance between their bases, (6) dorsum of promesonotum usually with <12 moderately coarse to coarse, irregular, longitudinal rugae that usually diverge toward humeral shoulders of pronotum or become transverse or rugoreticulate to vermiculate along anterior margin of pronotum, (7) in frontal view, frontal lobes and part to most of medial portion of clypeus and clypeal lobes usually blackish to black, notably darker than rest of cephalic dorsum, and (8) (HW <1.70 mm; ML <2.05 mm) ( Figure 106 View FIGURE 106 ).
Measurements — holotype (n = 12). HL 1.49 (1.40–1.63); HW 1.61 (1.42–1.69); MOD 0.35 (0.30–0.37); OMD 0.35 (0.32–0.41); SL 1.27 (1.04–1.35); PNW 1.12 (0.95–1.12); HFL 1.53 (1.37–.61); ML 1.82 (1.52–2.01); PW 0.45 (0.37–0.47); PPW postpetiole missing (0.50–0.61). Indices: SI 78.88 (65.09–80.50); CI 108.05 (99.34–106.96); OI 21.74 (18.34–24.03); HFI 95.03 (86.39–103.90).
Redescription. Head subquadrate to quadrate (CI = 99.34–106.96), widest just posterior to eyes; posterior margin flat to weakly convex in full-face view. Longitudinal rugae on cephalic dorsum prominent, in full-face view, medial rugae weakly diverging toward posterior corners of head. Interrugae on cephalic dorsum moderately to strongly granulate, dull; posterior corners rugose, interrugae smooth and shining to weakly granulate, weakly shining. Anterior margin of clypeus strongly concave, medial portion incised to depth of lateral lobes, dorsum with numerous subparallel, longitudinal rugae. Numerous long, curved, bristle-like, cream to light yellowish macrochaetae project from anterior margin of clypeus and basolateral margin of mandibles. Mandibles with six teeth; mandibular dorsum coarsely rugose. MOD ranging from 0.19–0.25× HL. In profile, eyes situated slightly anterior to middle of head, OMD = 0.91–1.23× MOD. In full-face view, eyes protruding slightly beyond lateral margins of head. Antennal scapes (SI = 65.09–80.50) failing to reach posterior corner of head by less than length of basal funicular segment; weakly striate longitudinally, interstriae weakly shining to shining; basal flange well developed, flattened with carinate margin. Psammophore well developed.
Mesosomal profile moderately convex; all mesosomal surfaces with prominent subparallel, irregular rugae, occasionally rugoreticulate on pronotal sides. In dorsal view, humeral shoulders of pronotum enlarged, dorsolateral margins weakly to moderately angulate. Dorsum of promesonotum with <12 moderately coarse to coarse, irregular, longitudinal rugae that usually diverge toward humeral shoulder of pronotum or become transverse or rugoreticulate to vermiculate along anterior margin of pronotum. Irregular rugae on pronotal sides traverse ventrally or posteroventrally or rugoreticulate; rugae on mesopleura angle posterodorsally; irregular transverse rugae on dorsum of propodeum traverse ventrally or anteroventrally on sides. Promesonotal suture often weakly to moderately impressed. Superior propodeal spines long, length> 0.6–0.8× the distance between their bases, spines connected by well defined keel; inferior propodeal spines slightly wider than high, apex weakly blunt to acuminate. Propodeal spiracles narrowly ovate facing posterad. Interrugae on mesosoma smooth and shining to strongly granulate, dull to weakly shining. Legs smooth and shining to weakly coriarious, weakly shining to shining.
Peduncle of petiole about as long as petiolar node, anteroventral margin with a well developed, rounded process. In profile, posterior surface of petiolar node weakly convex; node asymmetrical with anterior surface notably shorter than posterior surface, apex subangulate to angulate. In dorsal view, petiolar node longer than wide, widest near spatulate anterior margin, gradually narrowing posterad. Posterior surface of petiolar node with regular to irregular transverse or oblique rugae. Dorsum of postpetiole convex in profile; in dorsal view, widest at/near posterior margin, narrowing to anterior margin, maximum width and length about equal. Transverse rugae on dorsum of postpetiole finer, denser than those on posterior surface of petiolar node; interrugae on posterior surface of petiolar node and dorsum of postpetiole smooth and shining to weakly coriarious, weakly shining. First gastral tergum variable, most commonly smooth and shining to weakly or moderately coriarious at or near the base, weakly shining (62 of 71; 87.3%), with striae at base (7 of 71; 9.9%), or with striae covering up to anterior one-half of tergum (2 of 71; 2.8%).
Erect, short to medium-length, white pilosity moderately abundant on head, longest occasionally approaching MOD. Moderately abundant suberect to semidecumbent pilosity on scape; abundant decumbent hairs on funicular segments. Legs with moderately abundant suberect to decumbent white setae. Mesosoma, petiolar node, postpetiole, gastral terga with moderately dense, erect, white setae, longest hairs on mesosoma and petiolar node. Head dark reddish-brown to reddish-black; antenna, frontal lobes, mandibles, medial portion of clypeus blackish to black; rest of body black except for occasional reddish infusion on dorsum of promesonotum ( Figure 106 View FIGURE 106 ).
Queen diagnosis. Queens of this species are diagnosed by the following combination of features: (1) castespecific morphology of the mesosoma related to wing-bearing and presence of ocelli on head, (2) striae absent on first gastral tergum or present along base to sometimes extending over anterior one-third to one-half of segment, medioposterior one-quarter to one-third or more of first gastral tergum (including posterolateral margins) smooth and shining, (3) interrugae on cephalic dorsum weakly to moderately granulate-punctate, weakly shining to shining, (4) head reddish-brown to reddish-orange, mesosoma and gaster usually black, (5) pronotal sides concolorous blackish to black, anepisternum black, (6) in posterodorsal view, posterior corners smooth and shining to rugose, interrugae smooth and shining, (7) medial rugae along posterior margin of head regular to irregular, and (8) transverse or oblique rugae cover posterior surface of petiolar node and dorsum of postpetiole ( Figure 107 View FIGURE 107 ).
Measurements —(n = 13). HL 1.42–1.83; HW 1.59–1.87; MOD 0.34–0.41; OMD 0.34–0.47; SL 1.16–1.41; PNW 1.24–1.50; HFL 1.47–1.79; ML 2.19–3.01; PW 0.47–0.61; PPW 0.69–0.84. Indices: SI 65.19–78.86; CI 100.55–117.53; OI 18.58–23.27; HFI 85.25–98.16.
Description. With caste-specific morphology of the mesosoma related to wing-bearing and presence of ocelli on head. In full-face view, head quadrate to wider than long (CI = 100.55–117.53), widest just posterior to eyes, posterior margin flat to weakly convex. Longitudinal rugae on cephalic dorsum prominent, wavy; in full-face view, medial rugae diverging toward posterior corners of head; medial rugae along posterior margin of head regular to irregular; interrugae on cephalic dorsum weakly to moderately granulate-punctate, weakly shining to shining; posterior corners smooth and shining to rugose, interrugae smooth and shining. Mandibles with six teeth, dorsal surface coarsely rugose. Psammophore well developed.
All mesosomal surfaces except pronotal sides with subparallel, regular to wavy rugae, rugae irregular to rugoreticulate on pronotal sides, interrugae smooth and shining to weakly granulate-punctate, weakly shining; superior propodeal spines range from short, triangular teeth to moderately long, acuminate spines; inferior propodeal spines wider than tall, apex broadly rounded to subangulate. Peduncle of petiole long, anteroventral margin with weakly rounded process. In profile, petiolar node asymmetrical with anterior surface shorter than posterior surface, apex subangulate to angulate. Posterior surface of petiolar node with wavy, transverse to oblique rugae, interrugae weakly granulate-punctate, weakly shining. In dorsal view, postpetiole slightly wider than long; dorsum with wavy, transverse rugae that are finer, denser than those on posterior surface of petiolar node, interrugae weakly granulate-punctate, weakly shining. Striae absent on first gastral tergum or present along base to rarely extending over anterior one-third to one-half of segment, medioposterior one-quarter to one-third or more of first gastral tergum (including posterolateral margins) smooth and shining. Most body surfaces with moderately abundant suberect to erect, medium-length, whitish setae, one to two longer hairs along posterior margin of head approach MOD. Head reddish-brown to reddish-orange, rest of body black; mesoscutum, mesoscutellum, first gastral tergum sometimes with an orangish infusion ( Figure 107 View FIGURE 107 ).
Male diagnosis. Males of this species are diagnosed by the following combination of features: (1) first gastral tergum lacking striae, (2) body concolorous black to weakly bicolored, gaster sometimes slightly lighter than rest of body, but largely dark blackish orange to black, (3) erect hairs lacking on first gastral tergum, (4) in full face view, clypeus mostly to completely smooth and shining, polished, often with bluish to purplish reflections, and (5) notauli absent to faintly impressed ( Figure 108 View FIGURE 108 ).
Measurements —(n = 13). HL 1.10–1.29; HW 1.16–1.37; MOD 0.42–0.51; OMD 0.13–0.21; SL 0.41–0.52; HFL 1.45–1.81; ML 2.23–2.59; PW 0.48–0.61; PPW 0.68–0.82. Indices: SI 31.54–40.52; CI 98.35–114.66; OI 33.08–37.23; HFI 114.60–137.12.
Additional material examined. ARGENTINA: Chubut: 19.5 km E Shaman , 650 m, Nov 19, 1966 ( CASC) ; Esquel, Feb 19, 1949 ( IFML) ; 17.4 km S Dr Ricardo Rojas , 1870’, Jan 29, 2011 ( RAJC) ; Rt 26 at 2.3 km E Alto Río Mayo , 2050’, Jan 29, 2011 ( IFML; LACM; MACN; MCZC; RAJC; UCDC; USNM) ; Rt 259 at 23.9 km SW Tervelin , 1920’, Feb 22, 2014 ( RAJC) . Río Negro: Cerro Chall-huaco , 1805 m, Jan 2005 ( RAJC) ; El Bolsón, Feb 13, 1949 & Feb 20, 1971 ( IFML; LACM; MZSP) . Santa Cruz: 2.3 km S Cueva de las Manos, 2100’, Jan 31, 2011 ( RAJC) ; Ruta 40 at Estancia los Toldos (=Hostería Cueva de las Manos), Jan 8, 1999 ( IFML) ; 53.7 km S Jct Rt 40 & 29, 2080’, Feb 1, 2011 ( RAJC) ; El Calafete , 200 m, Jan 20, 1995 ( MCZ; UCDC) ; Rt 11 at 10.9 km E El Calafete , 830’, Feb 2, 2011 ( RAJC) ; Rt 11 at 19.3 km W El Calafete , 720’, Feb 2, 2011 ( RAJC) ; Rt 25 at 0.9 km E Tamil Aike , 1760’, Jan 31, 2011 ( RAJC) ; Rt 40 at 2.6 km N Bajo Caracoles , 1940’, Jan 30, 2011 ( RAJC) ; Rt 40 at 20.6 km S Bajo Caracoles , 2380’, Jan 31, 2011 ( RAJC) ; Rt 40 at 58.2 km S Bajo Caracoles , 2350’, Jan 31, 2011 ( RAJC) ; Rt 40 at 94.8 km S Bajo Caracoles , 2000’, Jan 31, 2011 ( RAJC) ; Rt 40 at 30.9 km S Perito Moreno , 1970’, Jan 30, 2011 ( IFML; RAJC) ; Rt 40 at 48.0 km S Perito Moreno, 1970’, Jan 30, 2011 ( RAJC) ; Rt 40 at 76.9 km S Perito Moreno , 2260’, Jan 30, 2011 ( RAJC) ; Rt 12 at 215.1 km SSW Pico Truncado , 2360’, Mar 10, 2017 ( RAJC) ; Meseta de la Vizcachas, Dec 1, 1943 ( IFML) ; no loc, no date (MACN). CHILE: Magallenes: Río Bagueles , Estancia Cerro Guido, Jan 7, 1955 ( IFML; LACM) ( Figure 91C View FIGURE 91 ) . Questionable locales (appear to be outside of geographic range): ARGENTINA: Chubut: Comodoro Rivadavia, no date ( MACN) . Santa Cruz: Rivadavia (= Comodoro Rivadavia?) (MACN) ( Figure 100C View FIGURE 100 ).
Etymology. In his description, Emery (1906) indicated that the mesosoma was covered with rugae that meander irregularly, hence the name vermiculatus (from Latin, vermiculatus = worm-shaped) (see also Kusnezov, 1951).
Discussion. Pogonomyrmex vermiculatus can be distinguished from other P. rastratus -group species based on: (1) first gastral tergum smooth and shining, striae rarely extending over more than anterior one-third of segment, (2) head dark reddish-brown to reddish-black, mesosoma and gaster black, and (3) superior propodeal spines long. Other P. rastratus -group species have striae that extend over more than one-half of the first gastral tergum and a different color pattern.
The identity of P. vermiculatus has been misunderstood since its description because of the paucity of verified material combined with the fact that no authors examined the holotype worker. It appears that Kusnezov (1951) examined only material from Meseta de las Vizcachas, Santa Cruz Province (IFML; #6756), and his description of P. vermiculatus followed that of Emery (1906), indicating that longitudinal striae were present along the base of the first gastral tergum with the rest of the segment smooth and shiny. Kusnezov (1951; 1978) also used “fine longitudinal striae along base of the first gastral tergum” to key out P. vermiculatus . I could not examine this character on the holotype worker because it was missing the postpetiole and gaster.
That Emery (1906) described P. vermiculatus based on one worker resulted in misunderstanding intra- and internidal variation in characters such as striae on the first gastral tergum. Collections made during this study demonstrate that workers display a range of character states within a colony and that the holotype worker of P. vermiculatus was not representative of the species because most workers lack striae along the base of the first gastral tergum (see above).
Confusion over the identity of P. vermiculatus continued into recent times when Snelling & Hunt (1975) synonymized three Chilean taxa, P. semistriata and P. intermedia , and P. spinolae under P. vermiculatus without knowing the actual identity of P. vermiculatus . In reality, P. vermiculatus appears to be rare in Chile where it is probably restricted to the extreme south; I have seen specimens from only Magdallenes Region.
Forel (1913) described P. vermiculatus var. chubutensis as a variety of P. vermiculatus because it differed from the type in: (1) lacking striae along the base of the first gastral tergum, and (2) rugae on the sides of the mesonotum traverse obliquely from posterodorsal to anteroventral while those on the pronotal sides traverse from the anterodorsal to posteroventral. As noted above, most workers of P. vermiculatus lack striae along the base of the first gastral tergum. Emery (1906) did not discuss patterning of rugae on the sides of the mesonotum and pronotal sides for the holotype of P. vermiculatus , but my examination of the holotype shows that rugae on the mesosoma traverse from anteroventral to posterodorsal while the pronotal sides are rugoreticulate. In the numerous series collected by the author, rugae on the pronotal sides are variable, and workers within a colony exhibit the range of patterns displayed by the holotypes of P. vermiculatus and P. vermiculatus var. chubutensis . Thus, I synonymize P. vermiculatus var. chubutensis under P. vermiculatus .
Lack of available material likely resulted in Forel (1915) describing P. vermiculatus var. jorgenseni from workers that were collected near the type locality of P. vermiculatus . Forel was apparently unaware of the proximity of these localities given that he compared P. vermiculatus var. jorgenseni to P. vermiculatus var. chubutensis , but not to P. vermiculatus . This omission may also have occurred because Forel assumed that striae along the base of the first gastral tergum were diagnostic for P. vermiculatus . Consequently, P. vermiculatus var. jorgenseni and P. vermiculatus var. chubutensis were compared because both forms were described as having a smooth and shining first gastral tergum (i.e., lacking striae near the base).
I examined 15 syntype workers of P. vermiculatus var. jorgenseni finding that the first gastral tergum varied in a manner similar to that described above for P. vermiculatus : the base of the first gastral tergum was smooth and shining on seven workers, weakly coriarious on six workers, weak longitudinal striae were present on one worker, and one worker had striae that extended over the anterior one-half of the segment.
Gallardo (1932) was the only author that directly compared P. vermiculatus and P. vermiculatus var. jorgenseni noting that the former form had punctures between the cephalic rugae whereas the latter form lacked depressed punctures between the cephalic rugae. This character also varied across syntypes of P. vermiculatus var. jorgenseni with punctures on the interrugae ranging from weak and weakly shining to lacking and smooth and shining. Thus, I synonymize P. vermiculatus var. jorgenseni under P. vermiculatus .
Biology. Workers of this species are solitary foragers. Nests of P. vermiculatus most commonly are placed in a clump of Stipa sp. (Poaceae) or at the edge of a rock and occasionally in an open, exposed site. The nest entrance ranges from a hole in the ground to a tumulus up to 15 cm in diameter; an external chaff midden is present on rare occasions. Nests are most easily located by baiting workers with cookie crumbs, then following them back to the nest. Partial nest excavations indicated that colonies contain up to approximately 2000 workers. Sexuals have been collected from January 29–February 22 indicating that mating flights occur during the austral summer. One intercaste also was collected ( Figures 3G–H View FIGURE 3 ). Several studies have examined foraging and recruitment biology of P. vermiculatus , but all of these studies probably used P. semistriata (see discussion under P. semistriata ).
Pogonomyrmex vermiculatus inhabits sites at elevations from 220–1805 m in the Valdivian temperate forests, Patagonian steppe, and Magellanic subpolar forests ecoregions, as defined by Olson et al. (2001) ( Figure 100C View FIGURE 100 ).





| LACM |
Natural History Museum of Los Angeles County |
| MACN |
Museo Argentino de Ciencias Naturales Bernardino Rivadavia |
| UCDC |
R. M. Bohart Museum of Entomology |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| MZSP |
Sao Paulo, Museu de Zoologia da Universidade de Sao Paulo |
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pogonomyrmex vermiculatus
| Johnson, Robert A. 2021 |
Pogonomyrmex vermiculatus var. jorgenseni
| Kusnezov, N. 1951: 253 |
| Gallardo, A. 1932: 141 |
| Forel, A. 1915: 353 |
Pogonomyrmex vermiculatus var. chubutensis
| Gallardo, A. 1932: 140 |
| Forel, A. 1913: 218 |
Pogonomyrmex vermiculatus
| Gallardo, A. 1932: 138 |
| Emery, C. 1906: 157 |