
Laonice (Appelloefia) barcinensis Sikorski, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4908.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:FD97747A-BF20-4820-B35E-67B432FF9987 |
|
DOI |
https://doi.org/10.5281/zenodo.4455622 |
|
persistent identifier |
https://treatment.plazi.org/id/03C687E2-FFF6-EE00-F0CB-5144DBF81C88 |
|
treatment provided by |
Plazi |
|
scientific name |
Laonice (Appelloefia) barcinensis Sikorski |
| status |
sp. nov. |
Laonice (Appelloefia) barcinensis Sikorski View in CoL sp. nov.
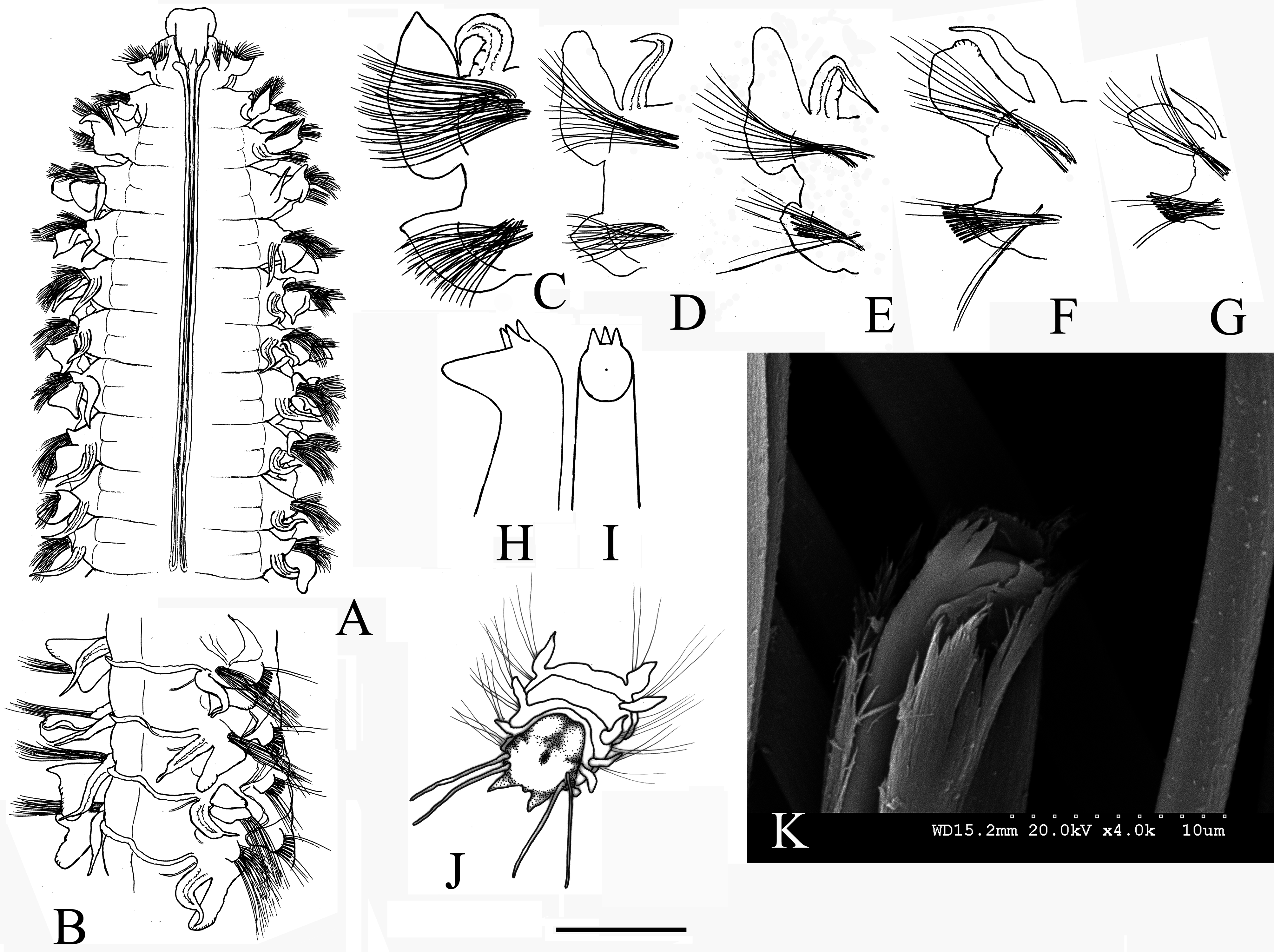
( Figures 3 View FIGURE 3 A–K, 4)
LSID: urn:lsid:zoobank.org:act:EC9FFD05-5EDF-46F7-A2C5-00874FCEC69C
Laonice bahusiensis — Sardá et al., 2009: 4, table 2. Material examined. Holotype ( MNCN 16.01 View Materials /18548): Foix submarine canyon, SW Barcelona, NW Mediterranean Sea , Projects EC-EUROMARGE-NB / CICYT-CONCENTRA, St. 05C21, 41.0333ºN 01.9333ºE, sediment trap moored 30 m above a sea ground 990 m deep, 04.1993–04.1994. GoogleMaps
Three paratypes from Foix submarine canyon, SW Barcelona, NW Mediterranean Sea, Projects EC-EURO-MARGE-NB / CICYT-CONCENTRA: St. 01C22, 40.9833ºN 02.0333ºE, sediment trap moored 30 m above a sea bed 1195 m deep, 04.1993–04.1994 ( MNCN 16.01 View Materials /18676); GoogleMaps St. 01C24 and St. 02C24, 41.0333ºN 01.9333ºE, sediment trap moored 500 m above a sea bed 990 m deep, 04.1993–04.1994 ( MNCN 16.01 View Materials /18677 and MNCN 16.01 View Materials /18617) GoogleMaps .
Description. Holotype complete, 126 chaetigers, 32 mm long, and 1.2 mm wide at about chaetiger 10. One paratype (MNCN 16.01/18676) complete—99 chaetigers, two other paratypes fragmented, however with pygidial fragments. Worms up to 61 mm long for 142 chaetigers, and 1.6 mm wide at chaetigers 7–12.
Prostomium longer than wide, from almost triangular, slightly T-shaped ( Fig. 3A View FIGURE 3 ) to almost quadrangular, with anterior margin truncated or slightly rounded with slight concavity, and completely separated from the peristomium at the anterior edges. Eyespots nearly invisible, smaller individuals may show one pair of faint (probably deeply embedded) eye-strips close to the base of the occipital antenna. Occipital antenna long and very thin (as long as or longer than notopodial postchaetal lamellae of chaetiger 1), ending in a thread-like tip, usually vertically erect, sometimes with middle bulbous enlargement, and inserted at the end of the prostomium. Intact palps lost from all specimens (one very short, torn off palp present in the sample MNCN 16.01/18617). Nuchal organs as long double U-shaped ciliary bands extending posteriorly to chaetigers 9–11 ( Fig. 3A View FIGURE 3 ). Anterior region of body (i.e. anterior segments bearing the nuchal organs plus the following one to approximately 10 anterior chaetigers) approximately 1.5 times wider than subsequent segments.
Branchiae from chaetiger 2, present nearly along the entire body, although absent in the 18–19 posterior-most chaetigers. Branchiae free, one third shorter than notopodial postchaetal lamellae until chaetiger 4, then gradually increasing in length and reaching about the same length as the notopodial postchaetal lamellae at chaetiger 7–9 ( Fig. 3A,C View FIGURE 3 ), and becoming 1.2–1.3 times longer than notopodial postchaetal lamellae on subsequent segments ( Fig. 3B View FIGURE 3 , D–F), except in the very posterior ones, where they become about one third shorter ( Fig. 3G View FIGURE 3 ). Branchiae from the mid-body ending in a threadlike process ( Fig. 3F View FIGURE 3 ), posterior-most branchiae thin, subulate ( Fig. 3G View FIGURE 3 ).
Notopodial prechaetal lamellae visible from chaetiger 3 to 12–22, attaining maximum size on chaetigers 9–10 (approx. 0.4 the length of notopodial postchaetal lamellae). Longest notopodial postchaetal lamellae at chaetigers 7–25. Notopodial postchaetal lamellae of the first 4–7 chaetigers leaf-like with elongated pointed tips ( Fig. 3C View FIGURE 3 ), more obtuse afterwards, with broadly rounded upper margins on chaetigers posterior to nuchal organs ( Fig. 3 View FIGURE 3 D–E), lacking an upwardly extension after chaetiger 45 ( Fig. 3F View FIGURE 3 ), becoming lanceolate at chaetiger 85 ( Fig. 3G View FIGURE 3 ) and finally disappearing 4 segments before pygidium.
Neuropodial prechaetal lamellae visible from chaetiger 2 to 12–13, reaching maximum size in chaetigers 7–9. Neuropodial postchaetal lamellae reaching their maximum size on chaetigers 8–17 ( Fig. 3C View FIGURE 3 ), then gradually diminishing further back, becoming very short on the posterior 40–45 chaetigers ( Fig. 3G View FIGURE 3 ). Neuropodial postchaetal lamellae bearing acute upwardly turned tips on the first 7 chaetigers ( Fig. 3C View FIGURE 3 ), with a peak gradually shifting along the margin of the lamella afterwards and becoming less evident ( Figs. 3 View FIGURE 3 D–E); neuropodial postchaetal lamellae with acute tips in the posterior half of body again ( Fig. 3 View FIGURE 3 F–G).
Transverse dorsal crests connecting bases of notopodial postchaetal lamellae ( Fig. 3B View FIGURE 3 ) from chaetiger 12–15 to chaetiger 50–74. Transverse dorsal crests generally low, attaining maximum height between chaetigers 15–47.
Inter-parapodial pouches from chaetigers 5–8 to nearly the end of the body, ending 5–7 chaetigers before pygidium. The inter-parapodial pouches may start from unpaired segments on opposite sides of the same specimen (difference of up to 2 segments).
Anterior parapodia with notopodial capillary chaetae arranged in more than 2 vertical rows in chaetigers 1–15 (with up to 6 rows in chaetigers 8–10), in 2 vertical rows from chaetiger 16; and neuropodial capillaries arranged in more than 2 vertical rows in the 12 anterior most chaetigers ( Fig. 3C View FIGURE 3 ), in 2 rows only on more posterior chaetigers. Neuropodial hooded hooks appearing from chaetiger 19–22, numbering 9–11 per bundle, and bearing 3 apical teeth above the main fang ( Fig. 3 View FIGURE 3 H–I, K). Sabre chaetae appearing from chaetiger 11–12, numbering at first 3–4 per bundle, and then 1–2 from chaetiger 16.
Pygidium with a pair of short ventral lobes and two pairs of thin and long, threadlike lateral anal cirri ( Fig. 3J View FIGURE 3 ).
No specific pigmentation observed.
Staining in Methyl Green. Anterior margin of prostomium and peristomium, occipital antenna, margins of postchaetal lamellae and small oval lateral spots between notopodia and neuropodia visibly staining more intensely than adjacent parts of body.
Etymology. The name barcinensis is derived from Barcino, ancient Latin name of Barcelona, and is given to this species in honor of the city of Barcelona, located close to the type locality of the new species.
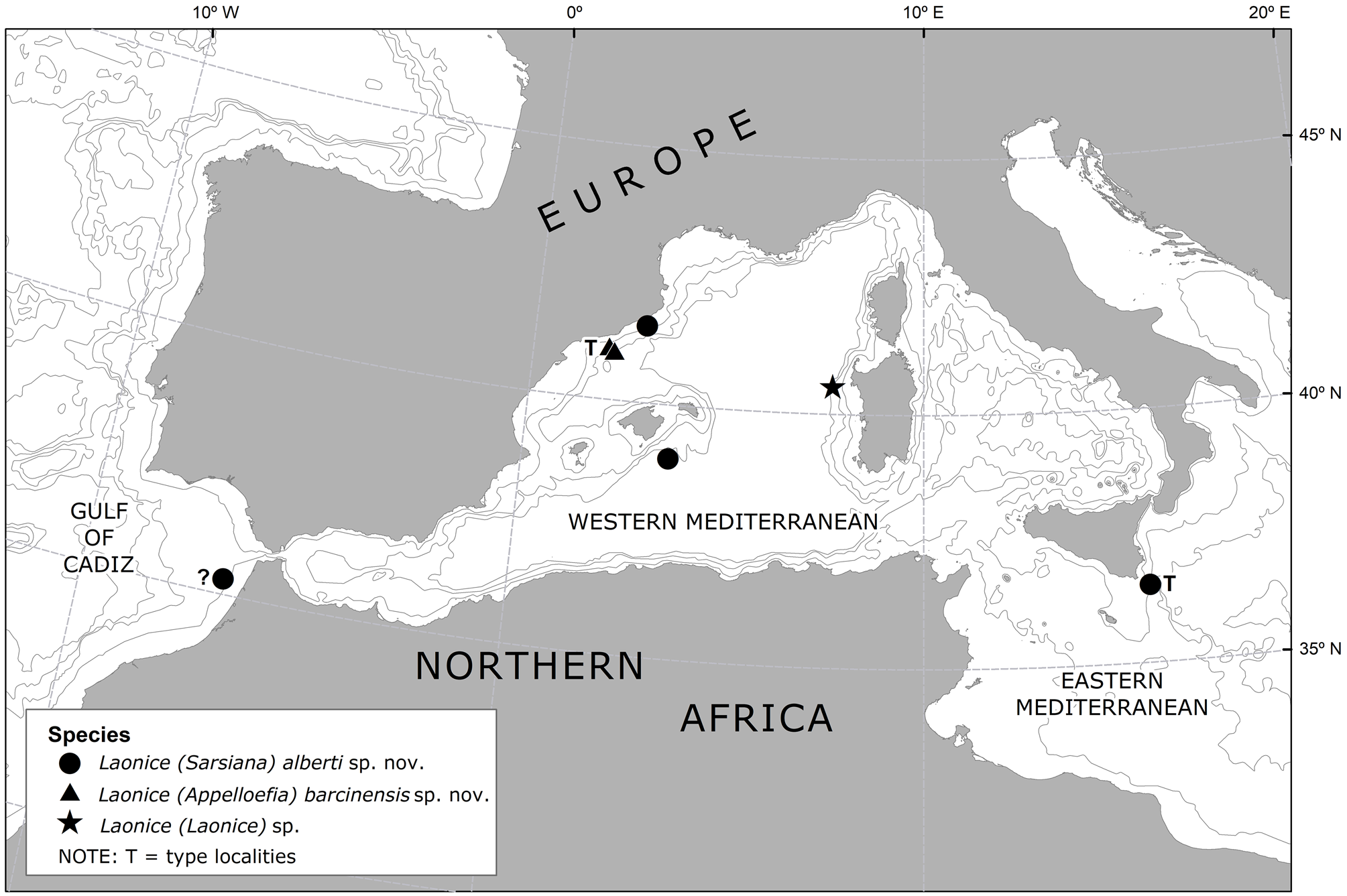
Distribution. Mediterranean Sea ( Fig. 4 View FIGURE 4 ): Foix submarine canyon (off NE Spain), in sediment traps moored 30 m above a seabed 990–1195 m deep, and 500 m above a seabed 990 m deep.
Remarks. Laonice (Appelloefia) barcinensis sp. nov. is morphologically very similar to L. (A.) appelloefi Ŝderstr̂m, 1920 and L. (A.) maciolekae Aguirrezabalaga & Ceberio, 2005 (synonymized with L. appelloefi in Meissner et al., 2014 ). However, branchiae and inter-parapodial pouches were only observed in L. appelloefi on a limited number of segments, whereas in the new species these structures are present through almost the entire length of the body, with the exception of the last posterior-most segments. These two characters suggest that the new species is similar to Laonice (Norgensia) norgensis Sikorski, 2003 , but this species has much longer nuchal organs, reaching chaetiger 15–29, and the inter-parapodial pouches appear from chaetiger 8–17 (against chaetiger 5–8 in L. barcinensis sp. nov.).
It is remarkable to notice how large adult worms of several dozens of milligrams of wet weight ( Sardá et al. 2009), belonging to a typically benthic genus, seem able to reach sediment traps moored as high as 500 m above the seabed. Sardá et al. (2009) hypothesized that the existence of a highly turbulent environment associated with the transport of large amounts of water and sediment inside the Mediterranean submarine canyons ( Canals et al. 2006), along with their strong bottom currents and resuspension events, would be responsible for the mobilization of sediment and benthic and epibenthic species into the water column. Such resuspension could even transport the specimens, up to hundreds of meters above the seabed where they normally live, reaching this way the sediment traps where they were found.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.