
Doleserpeton (Bolt & Lombard, 1985)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2008.00459.x |
|
persistent identifier |
https://treatment.plazi.org/id/73657737-FFBD-4B68-5B46-BB2CFED3CF0A |
|
treatment provided by |
Felipe |
|
scientific name |
Doleserpeton |
| status |
|
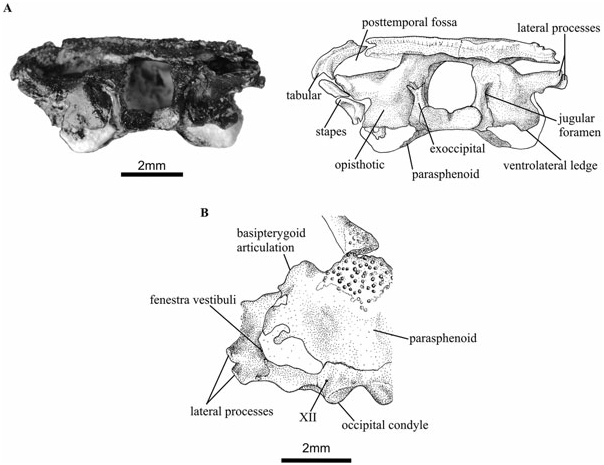
The distinct morphology and often highly ossified condition of many of the bony elements, including the inner ear, confirms Holmes’ (2000) assertion that Doleserpeton does not constitute a juvenile stage of other known dissorophoids. The otic capsules are large relative to those of other small temnospondyls, such as Micropholis ( Schoch & Rubidge, 2005) , and they constitute a dominant part of the occipital surface ( Fig. 5 View Figure 5 ). In disarticulated specimens, the otic elements are commonly found as separate prootics and opisthotics. However, in specimen UR1322 ( Fig. 5 View Figure 5 ), the two bones are fused without a discernable suture. In specimen UR1333, the exoccipital– basioccipital complex is fused to the opisthotic. The shape of the fenestra vestibuli is hard to study in detail, as it is damaged or partially obscured by other elements in the articulated specimens. It is bordered by the opisthotic, prootic, and, perhaps, the parasphenoid bones. It is unclear whether the parasphenoid constitutes its ventral edge, but it seems more likely that this area consisted of a cartilaginous part of the otic capsule proper. It appears to resemble the situation found in Eryops or Kamacops ( Schoch, 1999a,b), but the fenestra vestibuli of Doleserpeton is apparently relatively larger.
Prootic
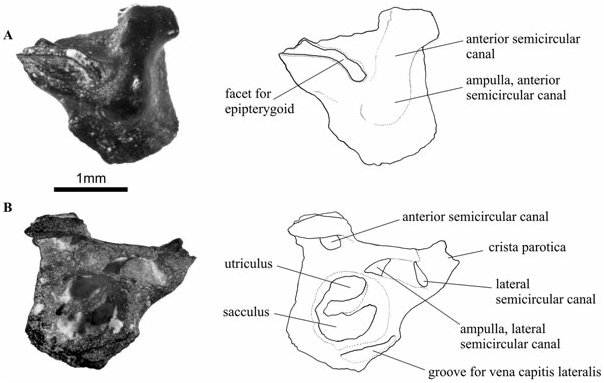
In anterior view ( Fig. 6A View Figure 6 ), the prootic is similar to that of Dendrerpeton , as described by Robinson et al. (2005). However, it lacks the groove for the internal jugular vein, with the anterior surface of the bone being smoothly convex. This convexity reflects the position of the anterior semicircular canal and its ampulla. The prootic has two raised articulating surfaces dorsally, separated by the anterior extension of the post-temporal fossa. The dorsolateral articulating surface has the appearance of a curved oblique groove on the dorsal and anterior surface of the crista parotica. The anterior portion of this groove contacted the posterior edge of the epipterygoid, or a cartilaginous extension of this bone. The dorsomedial articulation is a short elevated area that probably contacted the ventral surface of the parietal bone. The ventral area of the prootic contacts the parasphenoid– basisphenoid complex immediately above the basipterygoid articulation. A prootic was separated from the matrix of specimen UR1314, and this specimen reveals a great deal of detail of the anterior osseous labyrinth ( Fig. 6B View Figure 6 ). The plane of view cuts through the otic capsule immediately posterior to the level where the lateral semicircular canal joins its ampulla. The lateral and posterior semicircular canals are shown, as well as the ampulla of the lateral canal. The utriculus is separated ventrally from the compartment of the sacculus by a crescentic ossified wall, which only just fails to separate the two compartments completely. The overall configuration in this plane of view conforms very closely to the situation found in anurans, when viewing a transverse section through the otic capsule at the level of the ampulla of the lateral semicircular canal (e.g. Gaupp, 1904: fig. 159; T. Sigurdsen, pers. observ. of sections through the skull of Ascaphus ). Some of the anterior pockets of the perilymphatic system would presumably have been sited in this region of the otic capsule, situated dorsally in the space of the osseous labyrinth containing the utriculus, as well as an anterior diverticulum of the perilymphatic cistern bordering the sacculus ( Gaupp, 1904). However, no trace of an anteriorly situated perilymphatic duct (helicotrema) could be found in this part of the osseous labyrinth.
The ventrolateral groove in the prootic is in the position of the branch of the vena capitis lateralis (or ‘primitive head vein’), as seen in Ascaphus ( Pusey, 1943) . A more dorsal and medial branch of this vein was seen preserved as a result of calcification in the Tersomius specimen described by Carroll (1964).
Opisthotic
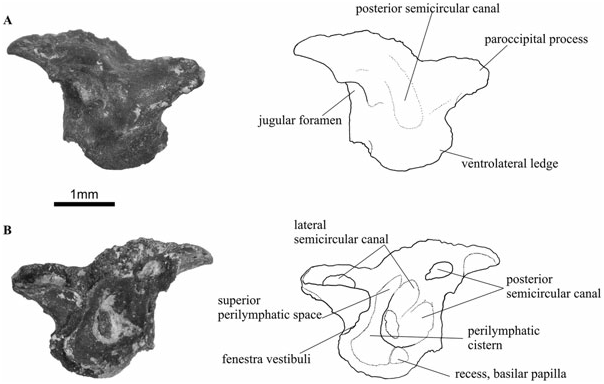
The opisthotic ( Figs 5 View Figure 5 , 7 View Figure 7 ) is a very large element of the braincase, as seen in occipital view. Three bony areas project from the main body of this element. These are the paroccipital process, located dorsolaterally, the medial ramus, situated above the exoccipital and foramen magnum, as well as the ventrolateral ledge, which is a bulbous area located ventral to the fenestra vestibuli. The medial ramus forms the roof of the foramen magnum, and it is fused with its fellow along the midline in specimen UR1322. The distinct shape of the otic capsule of Doleserpeton as compared with other temnospondyls is particularly noticeable because of the prominent ventrolateral ledge (the pseudobasal process of Wever, 1985). It is not fully ossified ventrally, but the position of the posteriormost parts of the parasphenoid may indicate its shape in life. The jugular foramen is visible between the opisthotic and exoccipital elements ( Figs 5A View Figure 5 , 7A View Figure 7 ).
In the well-preserved and mature specimen UR1322 the paroccipital process has two short lateral processes: one is located at the posterodorsal apex of the paroccipital process, whereas the other is situated slightly more ventrally and anteriorly ( Fig. 5A, B View Figure 5 ). The dorsal ridge contacted the tabular bone, and both ridges may have been attached to a tympanic annulus (see Discussion below). In posterior view, there is a convex area at the centre of the bone, immediately lateral and dorsal to the articulation with the exoccipital. This corresponds to the position of the posterior semicircular canal and its ampulla.
Internally, the best-preserved opisthotics (e.g. specimen UR1317) show several details of the posterior osseous labyrinth ( Fig. 7B View Figure 7 ). The plane of view cuts through the posterior and lateral semicircular canals twice, because of the curve of these canals at the posterior end of the membranous labyrinth. Anterior to the posteriormost portions of the semicircular canals, and positioned immediately medial to the fenestra vestibuli, is the perilymphatic cistern, of which part of the posterior wall has been preserved. A portion of the perilymphatic cistern ascends dorsally and medially to form the superior perilymphatic space. There is also a small indentation in the bone located medially and ventrally to the perilymphatic cistern. This probably marks the position of the recess for the basilar papilla.
The opisthotics show a slight ontogenetic range, in that the smaller specimens show little or no preservation of the lateral ridges, a less developed ventrolateral ledge, and medial rami of the opisthototic that are not fused along the midline. Thus, specimen UR1317 ( Fig. 7 View Figure 7 ) is apparently less mature than the only slightly larger specimen UR1322 ( Fig. 5 View Figure 5 ).
Exoccipital and basioccipital
The exoccipitals are fused to the basioccipital such that it is impossible to tell where one ends and the other begins. In mature specimens this complex is also partially fused dorsally to the medial ramus of the opisthotic, as in Edops ( Romer, 1942) .
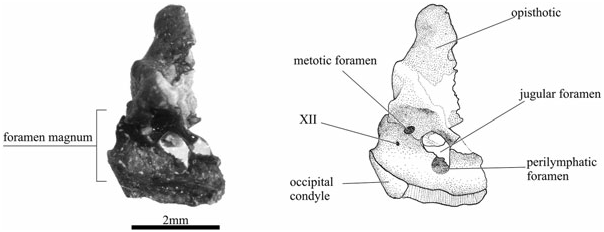
The exoccipital and basioccipital bones form the lateral and ventral walls of the foramen magnum ( Figs 5 View Figure 5 , 8 View Figure 8 ). Dorsally and laterally, the exoccipitals contact the opisthotics. The external jugular foramen opens between the two bones. The exoccipitals also form the two occipital condyles, which are slanted obliquely posterolaterally. A tiny foramen, probably for cranial nerve XII, pierces through the bone internally, half way between the foramen magnum and the fissura metotica, and opens externally immediately laterally to the condyle ( Figs 5 View Figure 5 , 8 View Figure 8 ).
In Doleserpeton , there are two or three foramina in the region of the fissura metotica, as seen in medial view ( Fig. 8 View Figure 8 ). The posterodorsal foramen, called the metotic foramen, is equivalent to a similar foramen found in Edops ( Romer, 1942) , Tersomius ( Carroll, 1964) , as well as in some amniotes (e.g. Rieppel, 1980), and it probably accommodated cranial nerves IX and X. Ventrally and anteriorly to this, there is a large foramen that is only rarely preserved in other temnospondyls. It is very similar to the anuran jugular foramen in shape and position, but it seems to be subdivided so that the anteroventral portion could be regarded as a separate foramen, the dividing wall possibly being finished in cartilage in life. This ventral part of the jugular foramen corresponds closely to the region where the perilymphatic duct communicates with the jugular foramen in modern anurans. It is therefore interpreted as the perilymphatic foramen. In the medial wall of the otic capsule, there are traces of the posterior acoustic foramen that would have been located immediately anterior to the perilymphatic foramen, but most of this region of the skull is missing.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |