
Crisicoccus seruratus ( Kanda 1933 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5209.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:78CF666B-8825-4B6F-93BE-86F892C2E1B4 |
|
DOI |
https://doi.org/10.5281/zenodo.7359552 |
|
persistent identifier |
https://treatment.plazi.org/id/03C687E9-FFB7-BF7F-A0F6-0C9CC93FFDD3 |
|
treatment provided by |
Plazi |
|
scientific name |
Crisicoccus seruratus ( Kanda 1933 ) |
| status |
|
Crisicoccus seruratus ( Kanda 1933) View in CoL
[Japanese common name: Matsumoto-kona-kaigaramushi]
Pseudococcus seruratus Kanda 1933: 133 View in CoL .
Pseudococcus matsumotoi Shiraiwa 1935: 66 View in CoL . syn. nov.
Pseudococcus astericola Shinji 1936: 49 View in CoL (synonymy by Kanda 1941: 24).
Crisicoccus seruratus ( Kanda 1933) View in CoL ; Paik 1978: 186 (change of combination); Kawai 2003: 314.
Crisicoccus matsumotoi ( Shiraiwa 1935) View in CoL ; Kawai 1980: 107 (change of combination).
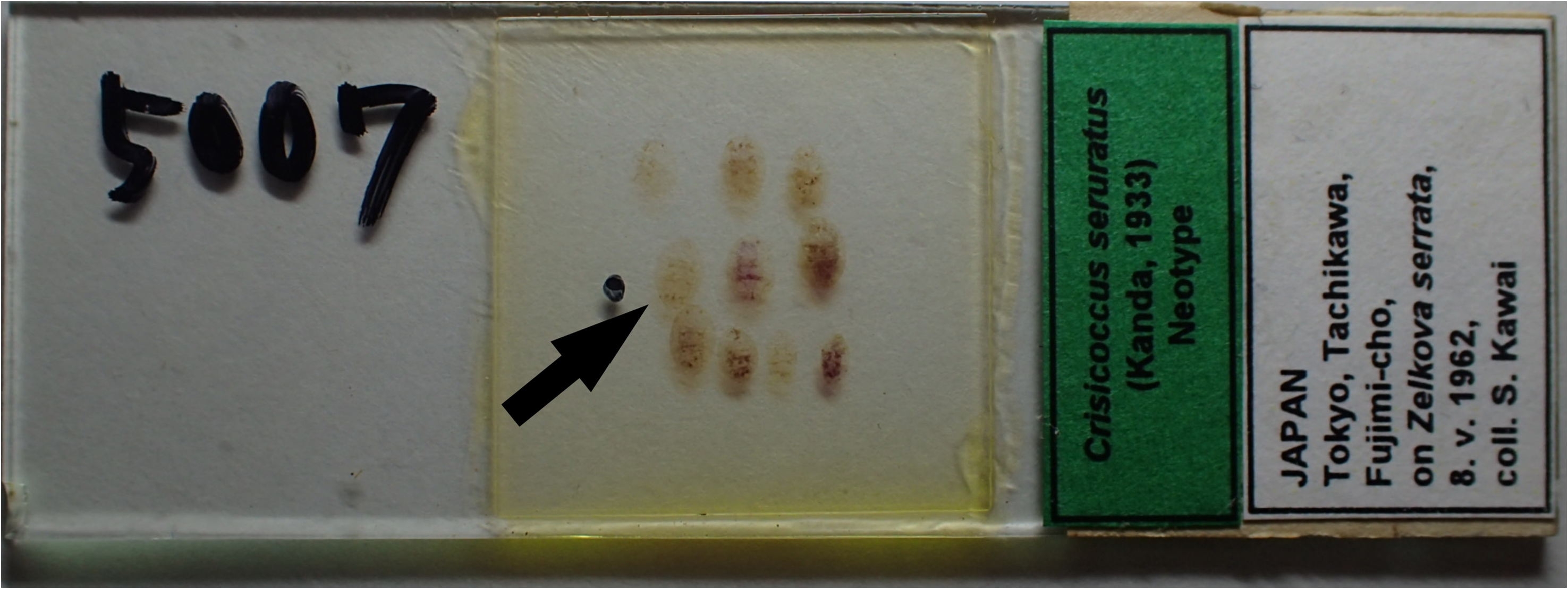
Material examined. Neotype, here designated: JAPAN / Tokyo, Tachikawa, / Fujimi-cho , / on Zelkova serrata , / 8.v.1962, / coll. S. Kawai; an adult female mounted with other 9 adult females on a slide ( KTUA). The individual at the left end of the middle row of specimens was selected as the neotype and is clearly indicated on the slide with a black ink dot ( Fig. 1 View FIGURE 1 ).
Other material. JAPAN: Tokyo, Chuo-ku, Hamarikyuteien , on Mallotus japonicus , 15. v.1972, coll. S. Kawai, 6 adult females mounted on 2 slides ( KTUA) ; Shizuoka prefecture, Shimizu, Okitsu , on Diospyros kaki , 8.vii.1976, coll. S. Kawai, an adult female mounted singly ( KTUA) ; Shimane prefecture, Izumo , on Vitis sp. , 25.v.2001, coll. S. Narai, an adult female mounted singly ( KTUA) ; Ibaraki prefecture, Kasama, Ago , on Pyrus pyrifolia var. culta , 27.iv.2022, coll. T. Tsunoda, 6 adult females mounted singly (3 ELKU, 3 EUMJ) .
Updated description
Appearance in life. Adult female 3‒4 mm long, dark purple covered with a white powdery wax. Projections of the wax secretion from body margin not so developed and limited to a few segments of posterior part of body. The body contents of this species turn blue black to dark green in 10% potassium hydroxide solution ( Kawai 1980, translated by HT).
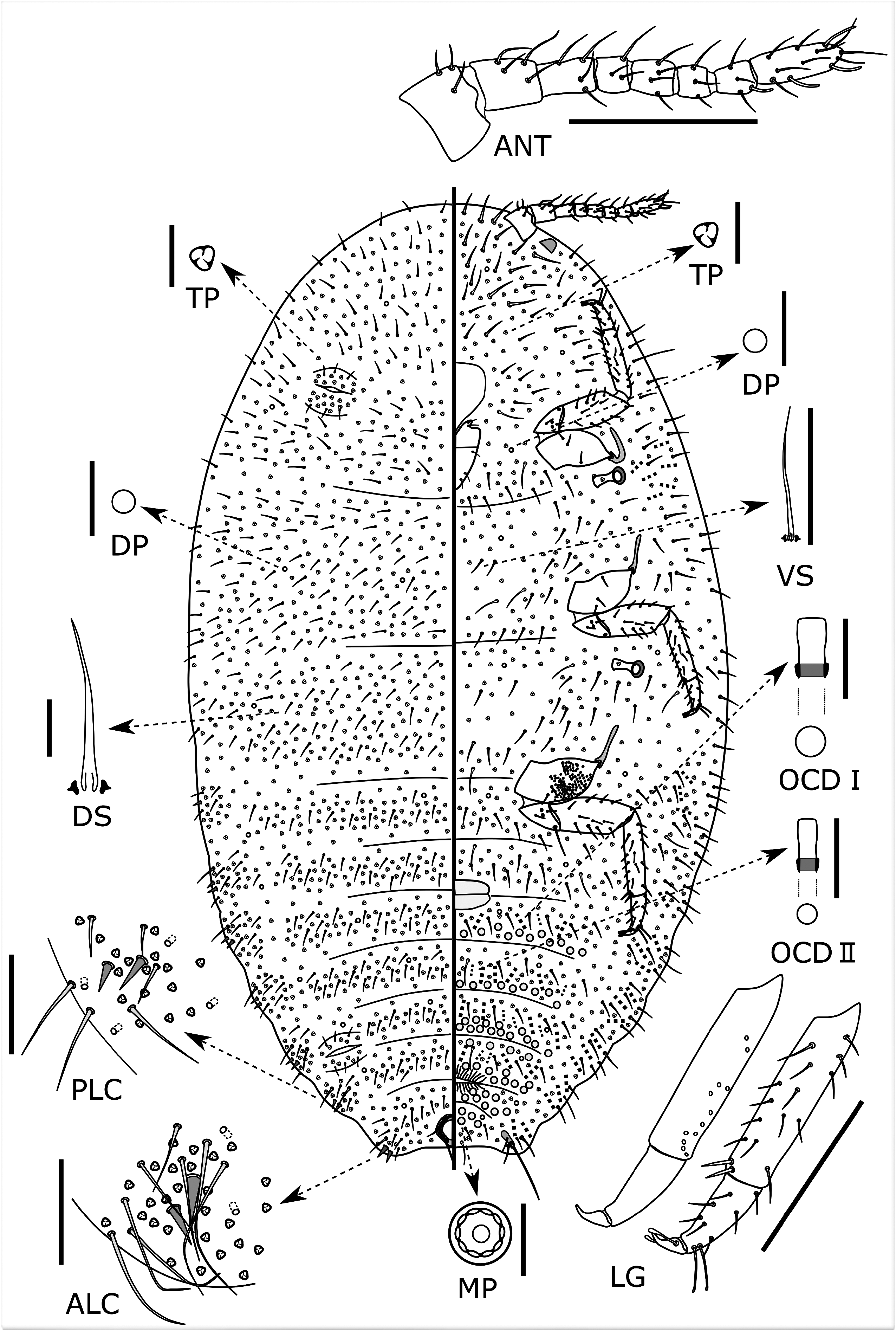
Slide-mounted adult female ( Fig. 2 View FIGURE 2 ) (n = 15). Body elongate oval, 3.6 (1.9–4.5) mm long and 2.2 (1.2–3.0) mm wide; derm membranous; segmentation recognizable, but not well developed. Anal lobes well developed, each with a long apical seta, 188–242 (145–270) µm long. Each lobe with a narrow and slightly faint anal lobe bar on ventral surface, but bar sometimes difficult to see in some specimens. Antenna 368–369 (368–500) µm long, with 8 segments and many flagellate setae; subapical segment with 1 fleshy seta and apical segment with 3 fleshy setae. Eyespot present on margin, not associated with discoidal pores. Legs well developed, with many flagellate setae; hind leg measurements (in µm): trochanter + femur 269–270 (269–394) long; hind tibia + tarsus 281–286 (281–377); claw 39–42 (34–45), without a denticle. Ratio of lengths of hind tibia + tarsus to trochanter + femur about 1: 1.04–1.06 (1: 0.89–1.06); ratio of lengths of hind tibia to tarsus 1: 1.78–1.83 (1: 1.78–2.60). Paired setose tarsal digitules present, subequal in length to the minutely knobbed claw digitules. Hind coxa with translucent pores on both surfaces; hind tibia usually with translucent pores on posterior surface; hind trochanter, femur and tarsus without translucent pores. Labium 170 (127–185) µm long, mostly shorter than clypeolabral shield. Circulus oval to quadrate, located between abdominal segments III and IV and divided by an intersegmental line, 70 (50–170) µm long and 90 (48–165) µm wide. Ostioles present, each with inner edges of lips weakly sclerotized; anterior ostioles each with a total for both lips of 22–27 (11–59) trilocular pores and 5 (2–7) setae; each posterior ostiole with a total for both lips of 24–27 (19–62) trilocular pores and 5–8 (2–8) setae. Anal ring 94 (90–113) µm wide, bearing 6 (6–8) setae, each seta 112–145 (60–170) µm long. Cerarii usually numbering 4 or 5 (4–8) pairs, mostly present on posterior abdominal segments, rarely present also on thoracic segments. Anal lobe cerarii (C 18) each containing 2 conical cerarian setae, each seta 17–20 (9–23) µm long and about 7 (5–8) µm wide at base, 4–5 (4–8) auxiliary setae and a concentration of trilocular pores. Penultimate cerarii (C 17) each containing 2 conical setae, 3 (0–8) auxiliary setae and a few trilocular pores. Cerarii situated further forward generally each with 2 conical setae with a few trilocular pores and auxiliary setae.
Dorsum. Setae flagellate, each 17–70 µm long, distributed usually segmentally; longest setae present on head. Trilocular pores each 3–4 µm wide, evenly distributed. Oral rim tubular ducts and oral collar tubular ducts absent. Discoidal pores each about 2–3 (1–4) µm wide, sparsely distributed.
Venter. Setae relatively long and flagellate, each 32–90 (19–168) µm long; setae on head longest. Multilocular disc pores, each 6–8 (6–9) µm wide, present in medial areas of abdominal segments IV‒IX, arranged in single row on each posterior area of abdominal segments IV‒V, in 2 or 3 rows on each posterior area of abdominal segments VI–VII, and in 1–2 rows on anterior areas of abdominal segments VI–VII but arranged randomly on abdominal segments VIII and IX. Trilocular pores, each same size as those on dorsum, evenly distributed. Oral collar tubular ducts of 2 sizes present: (i) large-type ducts, each about 3–4 µm in diameter, mostly wider than a trilocular pore, present on marginal to submarginal areas of all abdominal and thoracic segments and on medial areas of abdominal segments IV– VII, also forming transverse bands across abdominal segments IV–VII; and (ii) small-type ducts, each about 2–3 μm in diameter, mostly smaller than a trilocular pore, present on medial areas of abdominal segments IV–VII intermixed with large-type ducts, forming transverse bands across abdominal segments IV–VII. Discoidal pores, same size as those on dorsum, sparsely present.
Host plants in Japan. Buxaceae : Buxus microphylla ( Kawai 1980) ; Cannabaceae : Celtis sinensis ( Kawai 1980) ; Ebenaceae : Diospyros kaki ( Kawai 1980; Kawai 2003); Euphorbiaceae : Mallotus japonicus ( Kawai 1980) ; Fabaceae : Wisteria floribunda ( Kawai 2003) ; Juglandaceae : Juglans mandshurica ( Kawai 1980) ; Moraceae : Ficus carica ( Kawai 1980; Kawai 2003); Platanaceae : Platanus orientalis ( Kawai 1980) ; Rosaceae : Chaenomeles speciosa ( Kawai 1980) , Photinia glabra ( Kawai 2003) , Pyrus pyrifolia var. culta ( Kawai 1980; Kawai 2003); Sapindaceae : Acer palmatum ( Kawai 1980) , Acer spp. ( Kawai 2003) ; Ulmaceae : Zelkova serrata ( Kawai 1980) ; Vitaceae : Vitis spp. ( Kawai 2003) .
Remarks. The original description of C. seruratus by Kanda (1933) was based mostly on highly variable morphological characteristics (including proportions of antennal segment lengths) that have little taxonomic value. Most of Kanda’s specimens are deposited in Osaka Museum of Natural History, Japan (OMNH); however, a lengthy search was made for type specimens of this species in almost all of the Coccomorpha collections in Japan (including Osaka Museum of Natural History) but none could be found; we therefore conclude that all the type specimens of C. seruratus described by Kanda have been lost. The designation of a neotype of this species was necessary because the original description is not informative, complicated taxonomic problems are associated with the species, and the type material has been lost. One of the specimens used in this redescription is designated the neotype (above) for taxonomic stability. The neotype specimen was obtained from a tree of Zelkova serrata in the Kanto region of Japan, which is the same area where the primary type series for C. seruratus were collected. The morphology of this type specimen agrees well with the original description.
Currently, in Japan C. seruratus is recognized as C. matsumotoi . Its morphology differs significantly from “ C. matsumotoi ” reported by Ezzat & McConnell (1956) and Williams (2004), by having the following morphological characteristics (contrasting characteristics of the material incorrectly identified as “ C. matsumotoi ” are stated in parentheses): (i) a small number of cerarii, numbering fewer than 8 pairs (cerarii numbering 15–16 pairs); and (ii) oral collar tubular ducts of two different sizes present on venter (venter with only one size of oral collar tubular ducts). Therefore, in this study, we describe “ C. matsumotoi ” reported by Ezzat & McConnell (1956) and Williams (2004) as a distinct, new species, Crisicoccus ezzati sp. nov. (see below).
Crisicoccus seruratus has been collected on the roots of Zelkova serrata (= Abelicea serrata ), which is the host plant of Kanda’s original collection, but he used the name Abelicea serrata and he misspelt the scientific name of the host as “ Obelicea serurata ”. Kawai's (1980) view that C. matsumotoi and C. seruratus are the same species seems to be correct; however, he incorrectly designated C. seruratus (the older name) as a junior synonym of C. matsumotoi (García Morales et al. 2016) . Above, we formally synonymize Pseudococcus matsumotoi Siraiwa 1935 as a junior synonym of Pseudococcus seruratus Kanda. We were unable to examine any types of Pseudococcus astericola Shinji 1936 ; however, Kanda (1941) synonymized this species under Pseudococcus seruratus Kanda and we follow his synonymy.
In Korea, Paik (1978) and Kwon et al. (2003) regarded C. seruratus and “ C. matsumotoi ” as distinct species; however, the species they recognized as “ C. matsumotoi ” in Korea was a misidentification of Spilococcus pacificus (see below). Crisicoccus species do not have oral rim tubular ducts on the body surface, whereas Spilococcus species have at least a few so the genera can be easily distinguished from each other.
Crisicoccus seruratus shows similarities to C. melaleucae Williams 1985 (described from Queensland, Australia), such as possessing a small number of cerarii (≤ eight pairs), oral collar tubular ducts of two sizes, and lacking cerarii on the head; however, it differs from C. melaleucae in having relatively long dorsal setae, each approximately 17–70 µm long, whereas C. melaleucae has relatively short dorsal setae, each approximately 8–16 µm long.
| T |
Tavera, Department of Geology and Geophysics |
| EUMJ |
Ehime University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Crisicoccus seruratus ( Kanda 1933 )
| Tanaka, Hirotaka & Kamitani, Satoshi 2022 |
Crisicoccus matsumotoi ( Shiraiwa 1935 )
| Kawai, S. 1980: 107 |
Crisicoccus seruratus ( Kanda 1933 )
| Kawai, S. 2003: 314 |
| Paik, W. H. 1978: 186 |
Pseudococcus astericola
| Kanda, S. 1941: 24 |
| Shinji, O. 1936: 49 |
Pseudococcus matsumotoi
| Shiraiwa, H. 1935: 66 |
Pseudococcus seruratus Kanda 1933: 133
| Kanda, S. 1933: 133 |