
Centruroides hamadryas, Goodman & Prendini & Francke & Esposito, 2021
|
publication ID |
https://doi.org/ 10.1206/0003-0090.452.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5745655 |
|
persistent identifier |
https://treatment.plazi.org/id/510E87A5-6D7B-FFA0-25C5-FDA5FCA8F937 |
|
treatment provided by |
Carolina |
|
scientific name |
Centruroides hamadryas |
| status |
sp. nov. |
Centruroides hamadryas , sp. nov.
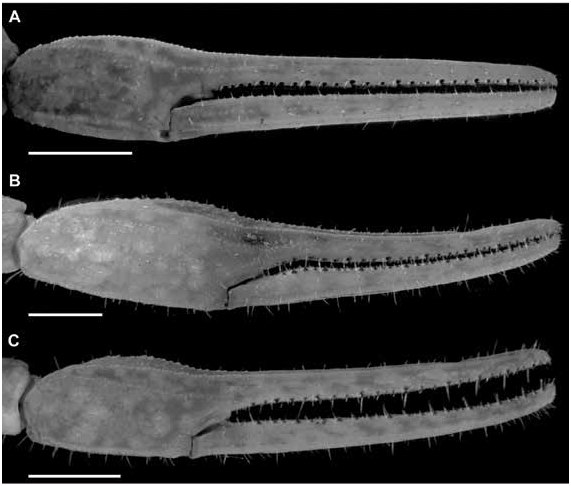
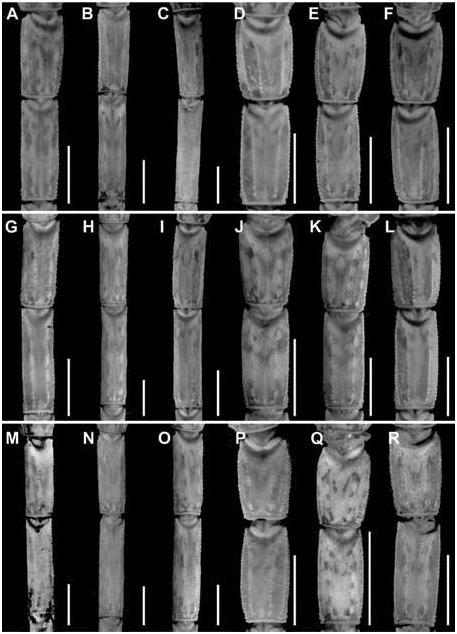
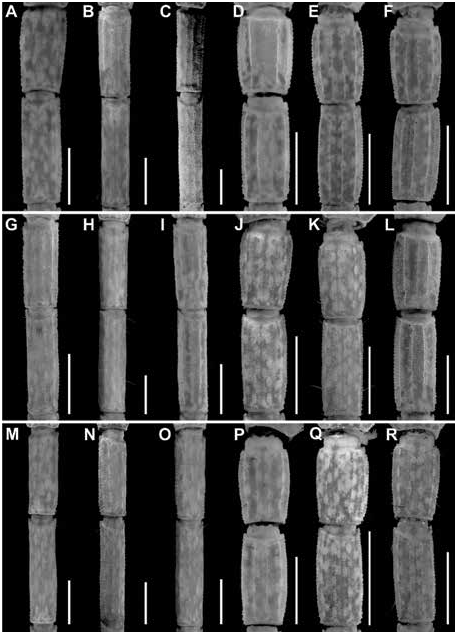
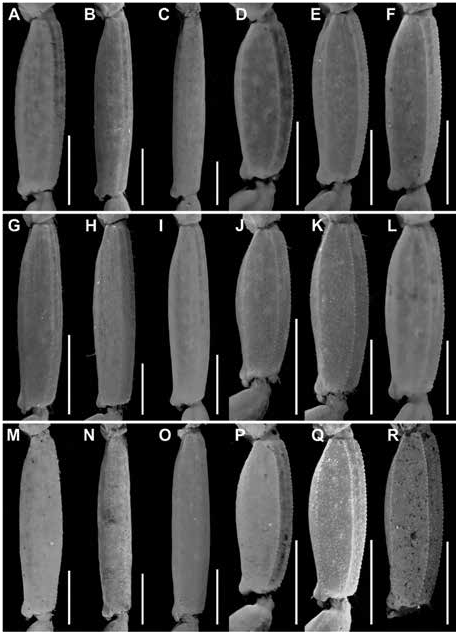
Figures 2 View FIGURE 2 , 4 View FIGURE 4 , 6A, B View FIGURE 6 , 9A, B View FIGURE 9 , 13A View FIGURE 13 , 14A View FIGURE 14 , 17B, E View FIGURE 17 ,
18B, E View FIGURE 18 , 19B, E View FIGURE 19 , 20B, E View FIGURE 20 , 21B, E View FIGURE 21 , 22B, E View FIGURE 22 , 23B, E View FIGURE 23 , 24B, E View FIGURE 24 , 25B, E View FIGURE 25 , 32 View FIGURE 32 , 33 View FIGURE 33 , tables 1 View TABLE 1 , 4 View TABLE 4 , 10 View TABLE 10
TYPE MATERIAL: MEXICO: Chiapas: Município Ocosingo: Holotype ♂ ( CNAN T01408 ), paratype ♂ ( CNAN T01412 ), paratype ♀ (CNAN T01413), La Galleta, 16°48′18.5″N 90°54′25″W, 103 m, 2.v.2005 A. Valdez, O.F. Francke, and A. Ballesteros, collected with UV light detection; paratype ♂ (CNAN T01414), paratype ♀ (CNAN T01415), same data, except: 2.v.1992, E. Barrera; 2 ♂ paratypes (CNAN T01409, T01411), paratype ♀ (CNAN T01410), same data, except: 114 m, 28.iv.2005, urban area toward blue water bridge.
ETYMOLOGY: The species name is noun in apposition, taken from the Greek nymph Hamadryas, mother of the hamadryads, tree-dwelling nymphs with lifelong bonds to the trees.
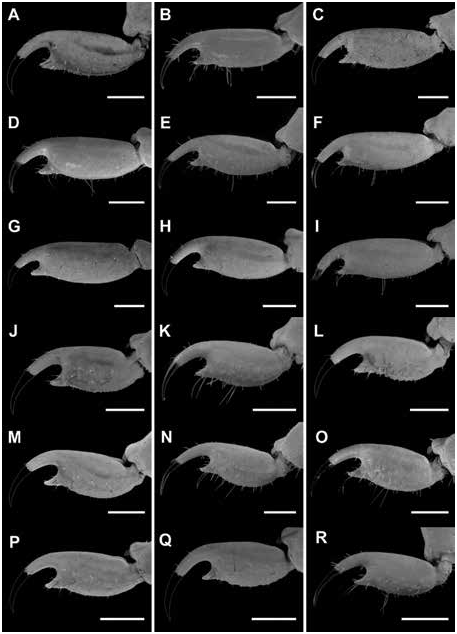
DIAGNOSIS: Centruroides hamadryas is most closely related to C. berstoni , from which it differs in the following respects. The carapace is densely granular, with distinct lateral ocular carinae, in the female of C. hamadryas (fig. 6B) but sparsely granular, more densely so on the interocular triangle, in the female of C. berstoni (fig. 6D). The pedipalp chela manus of the male is proportionally more incrassate in C. hamadryas (fig. 13A) than C. berstoni (fig. 13B). The legs of the male are less than 2× the length of the carapace in C. hamadryas but greater than 2× the length of the carapace in C. berstoni ( table 10 View TABLE 10 ). The dorsomedian carinae of the mesosomal tergites are distinct and complete in C. hamadryas but weakly developed and restricted to the posterior half of the segments in C. berstoni . The ventrolateral and ventrosubmedian carinae of mesosomal sternite VII are weakly developed to absent in C. hamadryas , whereas the ventrolateral carinae are weakly developed and the ventrosubmedian carinae absent in C. berstoni . The telson vesicle is densely setose in C. hamadryas but sparsely setose in C. berstoni .
Additional differences between C. hamadryas and other species of the clade are as follows. The retrodorsal carina of the chela manus is complete and the dorsomedian carina weakly developed and restricted to the distal half, in the male of C. hamadryas (fig. 13A), whereas the retrodorsal carina is weakly granular and the dorsomedian carina absent in the male of C. berstoni (fig. 13B), C. catemacoensis (fig. 11B), C. cuauhmapan (fig. 11C), and C. rileyi (fig. 11A). The retrodorsal carina of the manus of the female is complete and the prodorsal carina restricted to the distal third in the female of C. hamadryas (fig. 13B), whereas the retrodorsal carina is finely granular and the prodorsal carina absent in the female of C. berstoni (fig. 14B), C. catemacoensis (fig. 12B), C. cuauhmapan (fig. 12C), and C. rileyi (fig. 12A). The pedipalp chela fingers bear short, dense setation in C. hamadryas but sparse setation in C. berstoni . The dorsosubmedian and dorsolateral carinae of metasomal segments I–III are well developed and granular in the male of C. hamadryas (figs. 17–19E) but weakly developed to absent in the male of C. berstoni (figs. 17–19K). The telson vesicle of the male is not posteriorly bilobed in C. hamadryas (figs. 23–25B), unlike C. C. cuauhmapan (figs. 23–25G) and C. rileyi (figs. 23–25A). The vesicle of the female is shorter and more robust, with intercarinal surfaces smooth in C. hamadryas (figs. 23–25E), but sparsely granular in C. berstoni (figs. 23–25K) and densely granular in C. catemacoensis (figs. 23–25P), C. cuauhmapan (figs. 23–25J), and C. rileyi (figs. 23–25D).
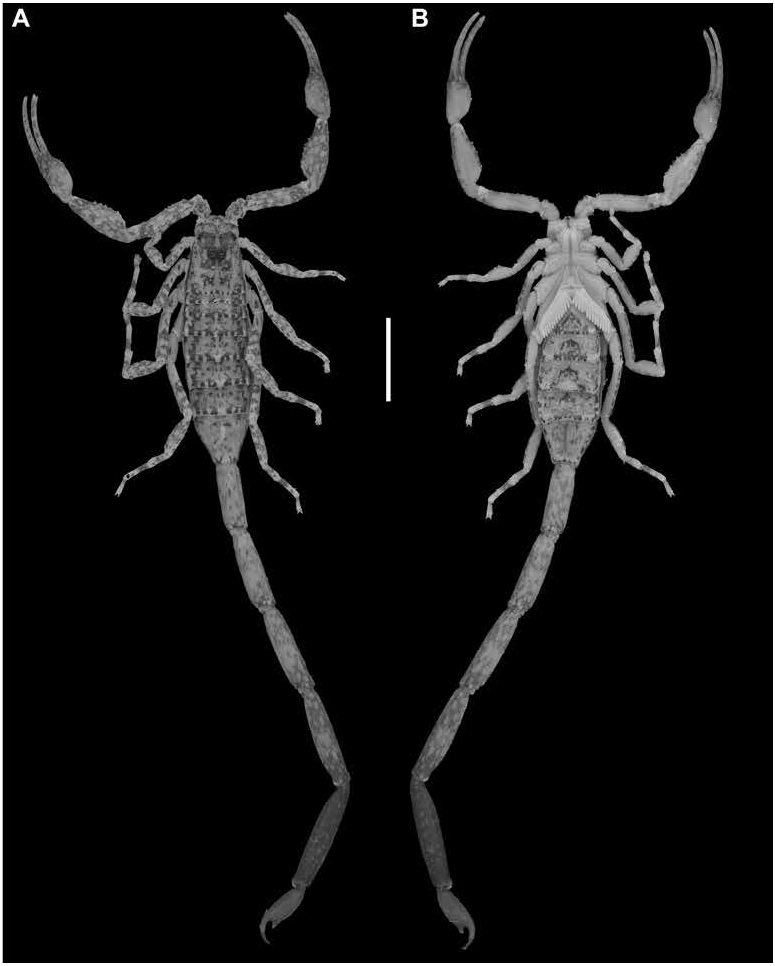
DESCRIPTION: The following description is based on the holotype male, with differences among other material noted in the section on variation.
Coloration: Base color pale yellow, with extensive infuscation, creating mottled or marbled pattern. Carapace with uniformly infuscate marbling, more densely infuscate medially. Pedipalp chela fingers and manus, dorsal and retrolateral intercarinal surfaces with moderately infuscate marbling; prolateral and ventral intercarinal surfaces mostly immaculate. Legs retrolateral surfaces with infuscate marbling; prolateral surfaces pale, immaculate. Tergites with unformly infuscate mottling, pale stripe medially, blackish spots submedially, and faint, narrow bands laterally. Sternites pale, with faintly infuscate triangular to trapezoidal marking at posterior margin of sternite III, fading to infuscate mottling on sternite VII. Metasomal segments uniformly, faintly marbled; segment V and telson markedly infuscate, noticeably darker than preceding segments.
Carapace: Shape trapezoidal; anterior width four-fifths of posterior width ( table 4 View TABLE 4 ); anteromedian sulcus moderately deep, oval; posteromedian sulcus shallow anteriorly, deeper posteriorly; median ocular tubercle moderately granular; carinae weakly developed, comprising small to medium-sized granules (fig. 6A).
Pedipalps: Orthobothriotaxic, Type A; femur dorsal trichobothria with α configuration; pedipalp chela fixed finger, trichobothrium db situated slightly distal to et. Femoral carinae serrate; retromedian carinae comprising spiniform granules; dorsal intercarinal surface moderately granular; prolateral surface with series of large spiniform granules. Patella prodorsal, dorsomedian, retrodorsal and proventral carinae moderately developed, serrate; retromedian carina well developed, serrate; retroventral carina incomplete, serrate; prolateral intercarinal surface with five or six large, subspiniform granules. Chela manus proventral carina moderately developed, comprising few rounded granules; other carinae weakly developed, granular. Fixed finger, median denticle row comprising eight oblique subrows, each flanked by pro- and retrolateral supernumerary denticles. Movable finger, median denticle row with short terminal row comprising four denticles preceded by eight oblique subrows, each flanked by pro- and retrolateral supernumerary denticles.
Legs: Leg I length 1.88× greater than carapace length ( table 10 View TABLE 10 ). Telotarsi ventral surfaces densely covered with short setae; ungues markedly curved.
Pectines: Pectinal plate 1.9× wider than long; posterior margin distinctly rounded; pectinal tooth count 14/14 (♂) (fig. 6A, table 4 View TABLE 4 ).
Mesosoma: Tergites width similar to carapace posterior width; I and II slightly narrower ( table 4 View TABLE 4 ). Pretergites surfaces smooth to finely granular. Posttergites surfaces weakly granular; I–VI with dorsomedian carinae moderately granular; VII surface weakly granular, dorsomedian carina moderately granular, dorsosubmedian carinae serrate, dorsolateral carinae well developed. Sternites III–VI, surfaces smooth; VII surface weakly granular, ventrolateral carinae reduced to few granules.
Metasoma: Metasoma length 3.01× mesosoma length ( table 4 View TABLE 4 ). Segments longer than wide; increasing in length posteriorly, segment V 2× length of I; carinae complete, granular on segments I–III, other carinae absent or obsolete; intercarinal surfaces sparsely granular (figs. 17–22E).
Telson: Vesicle elongate, ovoid; ventral surface shallowly convex, sparsely granular posteriorly; ventromedian carina granular, terminating at subaculear tubercle; subaculear tubercle narrow and angular in lateral aspect, directed toward midpoint of aculeus. Aculeus angled ventrally at slightly less than 90° (fig. 25B, E).
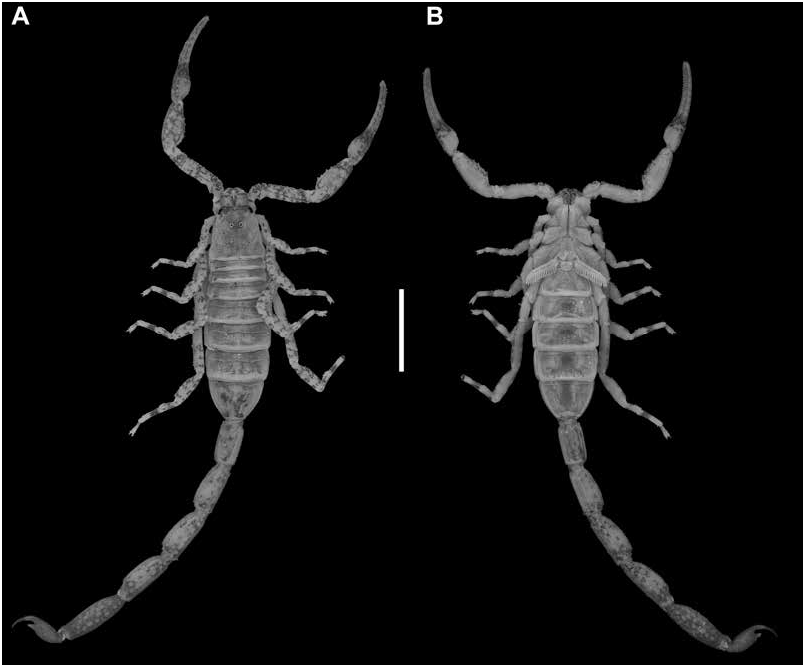
Variation: Adult males and females differ as follows. The pectinal tooth count is slightly higher (14 or 15), the mesosoma proportionally longer and slenderer, and the metasoma up to 3× longer, with segment V also roughly 1.5 mm longer, in males (figs. 23B, E, 24B, E, 25B, E, table 4 View TABLE 4 ). The tegument is more densely infuscate, the pectinal plate produced into a rounded lobe posteriorly, which is punctate and slightly infuscate, the pectinal tooth count slightly lower (12 or 13), and the telson shorter and narrower, in females (figs. 9A, B, 23B, E, 24B, E, 25B, E, 32A, B, 33A, B, table 4 View TABLE 4 ).
DISTRIBUTION: Centruroides hamadryas is known only from the state of Chiapas in southeastern Mexico, but may extend across the Usumacinta River into Guatemala. The known records occur in the Lacondón Forest, on the northern edge of the Montes Azules Biosphere Reserve (fig. 4).
ECOLOGY: The localities at which C. hamadryas has been recorded range in altitude from 103 to 153 m, all situated in a lowland tropical rainforest. Most specimens of this strictly arboreal species were located with UV light detection at night and captured by holding an insect net beneath the branch on which they were sitting and tapping the branch with a stick; their escape reaction is to drop immediately to the leaf litter below, where they invariably disappear. The habitat and habitus are consistent with the arboreal, corticolous ecomorphotype ( Prendini, 2001a).
REMARKS: Specimens from Frontera Corozal, Chiapas, Mexico, were misidentified as C. schmidti by Francke (2007). Teruel and Stockwell (2002) and Francke (2007) noted differences among the pectinal counts of specimens from Chiapas, with 13–16 (♂) and 13–14 (♀), and Honduras, with 12–15 (♂) and 13–15 (♀). Although this slight variation in pectinal tooth counts does not provide sufficient evidence to distinguish between C. hamadryas and C. schmidti , additional morphological differences, outlined in their respective diagnoses, together with genetic divergence among samples from the two areas, confirmed the distinction between them.
MATERIAL EXAMINED: MEXICO: Chiapas: Município Ocosingo: La Galleta, 2 km SE of Frontera Corozal, 16°48′12.7″N 90°52′11.1″W, 132–150 m, 28.iv.2004, R. Paredes and J.L. Castelo, collected with UV light detection, 2 ♂ (AMNH [LP 2948]), 1 ♀ (CNAN SC3987), 16°49′55″N 90°56′08″W, 146 m, 7.iv.2005, A. Valdez, O.F. Francke, and A. Ballesteros, collected at night with UV lamp, 1 juv. ♂ (CNAN SC3986), 16°48′18.5″N 90°54′25″W, 114 m, 28.iv.2005, A. Valdez, O.F. Francke, and A. Ballesteros, urban area toward blue water bridge, collected with UV light detection, 2 ♂, 1 ♀, 1 juv. ♂, 1 juv. (CNAN SC3988).










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.