
Arawakis poinari, Opitz, Weston, 2007
|
publication ID |
https://doi.org/ 10.1649/0010-065x(2007)61[339:apongn]2.0.co;2 |
|
persistent identifier |
https://treatment.plazi.org/id/87654039-FFAA-FFF0-B5BE-AB73FC2EFEDA |
|
treatment provided by |
Carolina |
|
scientific name |
Arawakis poinari |
| status |
sp. nov. |
Arawakis poinari , new species
( Figs. 1–3 View Figs View Fig )
Holotype: Female. Dominican Republic: Cordillera Septentrional, 1986, George Poinar, Jr. The holotype is embedded in a triangular piece of Dominican Amber with approximate side-length dimensions of 10 mm 3 8 mm 3 8 mm. The amber specimen has a wedge shape, is 2 mm at its greatest thickness, and has one polished and one roughened surface. It is maintained in a 30 mm 3 30 mm clear plastic box set on a cushion of cotton. The clear plastic box is labeled C, 7-72A (accession number). The specimen is deposited in the Poinar Collection held in the Entomological Collection of Oregon State University , 2046 Cordley Hall , Corvallis, Oregon 97331-2907.
Description: Size: Length 5.0 mm; width 1.8 mm. Integument: Cranium, prothorax, elytra, and legs pale brown, remainder dark brown. Head: Considerably wider than pronotum; eyes bulging, coarsely faceted, and only slightly emarginated; antenna comprised of eleven antennomeres, subfiliform, with loose antennal club ( Fig. 3 View Fig ), antennomere 11 about three times longer than wide; labial palpomere 3 securiform; maxillary palpomere 4 digitiform; gula broad. Thorax: Pronotum elongate, subscabrous, outer margins only slightly expanded at middle, anterior transverse depression feebly expressed, procoxal cavities open; pro- mesotarsus comprised of five tarsomeres; profemora notably swollen, meso- metafemora slightly swollen; mesotibia with two spurs; protarsus and mesotarsus with four pulvilli; mesotarsal claws with well-defined denticle; elytra elongate, serially punctate, slightly extended beyond distal end of abdomen. Abdomen: Venter comprised of six visible sterna; posterior margin of sixth visible sternum and pygidium evenly arcuate.
Etymologies: The generic name is a dedicatory name taken from Arawak, the name of an aborigine sector of Hispaniola, and the Latin patronymic suffix – is. The specific epithet is dedicated to George Poinar, Jr. for his many contributions to fossil research.
Evolutionary Relationships
The holotype presents a combination of characteristics that are most compatible with those of the extant genera of subfamily Clerinae . The coarsely facetted eyes, four metatarsal pulvilli, eleven antennomeres, and serially punctate elytra places this vanished species akin to a relatively primitive line of new world clerines now represented by the genera Priocera , Axina , and such Chilean genera as Natalis . A. poinari differs from these genera by its secondary loss of the pronotal disc vertical fissure and by its possession of an enlarged fourth tarsal pulvillus ( Fig. 3 View Fig ) which is apomorphic for the genus. The closest living relative of Arawakis appears to be the Chilean genus Natalis with which Arawakis shares an apomorphic characteristic of the antennae. The species of these two genera have an antennae in which antennomeres 3 to 8 become increasingly serrate and antennomeres 9 to 11 form a distinct club. In the abovementioned other genera antennomeres 3 to 10 are distinctly serrate. Antennal serration is also a characteristic of the North American genus Aulicus Spinola and the mostly Asiatic Tillicera Spinola. However , in specimens of these two genera the tarsal claws lack a basal denticle, which eliminates then from intergeneric relationship consideration.
The elongation of the eleventh antennomere, present in A. poinari , is reminiscent of the antennal condition present among the species of the African genus Phloiocopus Spinola. However , the similarity between these geographically widespread taxa probably represents an expression of homoplasy rather than a manifestation of synapomorphy. In the clerine genus Phloiocopus , the terminal maxillary palpomere is securiform and the tarsal claws lack basal denticles. In A. poinari , the terminal maxillary palpomere is clearly digitiform and the tarsal claws have well-developed basal denticles. Moreover, length of the eleventh antennomere has shown to be rather flexible genotypically as its distribution across subfamilial lines suggests. Its length is sex dimorphic in the tilline genera Lecontella Wolcott and Chapin and Cymatodera Gray with some males showing the terminal antennomere greatly extended. I suggest, therefore, that among the taxa in question the properties of the maxilla and tarsal claws are more reliable than the shape of the 11th antennomere. Therefore, I suggest that Arawakis may be more credibly aligned evolutionarily with the primitive, predominantly South American group of clerines and most closely related to the Chilean genus Natalis Laporte. I consider the serially arranged punctations on the elytra of A. poinari , and the South American stock as represented by Natalis and allies, synapomorphic for this hypothesized Greater Antilles-South American lineage.
As previously noted, the pronotal disc of the holotype of A. poinari does not show a ‘‘middiscal vertical fissure’’ a characteristic potentially valuable for predicting suprageneric relationships. This fissure is prominent in widespread Old World genera and is present among Priocera Kirby , Axina Kirby , and the Chilean group of clerines to which Natalis Laporte belongs. Its absence in A. poinari is considered a secondary loss.
Zoogeographical Considerations
The usefulness of insect amber fossils for stimulating discussions of ancient morphologies and evolutionary relationships, or for elucidating historical biogeography, is amply documented. See, for example, the works of Grimaldi (1992), Grimaldi, et al. (1993), Grimaldi and Amorim, D. S. (1995), Sturm and Mendes (1998), Poinar (1999 a, b), Manley and Poinar (1999), and references therein. In my view, the most significant use of amber fossils towards paleographic analyses is that the fossils provide a minimum age for the taxon that any particular fossil represents. Grimaldi (1992:179) points out, however, that such fossils may also indicate ‘‘phenotypic and/or geographical extinctions’’ that may shed some light on whether an extant distribution is based on vicariance biogeography or on geographic extinctions.
The geologic age of amber fossils is, of course, pivotal information to establish the minimal geologic age of a taxon which, in turn, can have an important bearing on the hypotheses proposed about the temporal history of the evolutionary lineage to which that taxon belongs. It is generally agreed that Baltic Amber is approximately Lower Eocene in age, some 40 million years ago, while Dominican amber has been dated from 15–45 million years ( Schlee 1990: 53; Grimaldi 1995 a: 210; Poinar 2000: 1040). Iturralde-Vinent and MacPhee (1996) provide convincing evidence and argumentation, based on biostratigraphy and paleogeographic data, that Dominican Amber was formed during the late Early Miocene through early Middle Miocene; some 15–20 million years ago. For a different viewpoint, the reader is referred to the comprehensive work of Poinar and Mastalerz (2000). This contribution provides a general review of most of the published works about the estimation of the age of Dominican amber and the taphonomy of fossilized resins.
Although application of anatomical features to the principle of ‘‘Occam’s Razor’’ excludes a possible close relationship between A. poinari , with the African taxon Phloiocopus Spinola , the literature does include Dominican fossil whose inclusions have Old World affinities ( Grimaldi 1991, 1995 b; Sturm and Mendes 1998; Poinar 1991).
The enlarged fourth tarsal pulvillus and the elongation of the terminal antennomere are two derived characteristics that define A. poinari . The increasingly serrate conditions of antennomeres 3 to 8 in conjunction with the presence of a definitive antennal club establishes a synapomorphic relationship between Arawakis , Natalis , and possibly the Nearctic Aulicus Spinola or the Asiatic Tillicera Spinola ; to the last two mentioned genera if we consider the basal denticle of the tarsal claw a homoplasic development. As the holotype specimen of A. poinari , is a female, convention has it that the male would have the last antennomere much more extended. This kind of sexual dimorphism of antennal structure is seen in the New World tilline genus Lecontella and the African clerine genus Phloiocopus .
The absence of the discal pronotal fissure, in A. poinari , suggests that considerable evolution had occurred from the historical divergence of Natalis and Arawakis , from the Natalis - Arawakis ancestor. The absence of the pronotal fissure in A. poinari may be explained as follows. Perhaps, the progenitor of A. poinari , involved an ancestor that separated very early from its South American stock, early enough to have lost the pronotal fissure. It is possible that the progenitor of the Central American Arawakis -like ancestor, was a derivative from a Gondwanian lineage that in the New World emigrated northward only to eventually succumb to the vicissitudinous climate and geology of the Cretaceous Central American archipelago (Malfait and Dinkelman 1972, fig. 3). The widespread cooling during the Plio-Pleistocene may have also been a significant factor in the disappearance of this ancient clerine lineage from Hispaniola and environs of Central America. Poinar (1999 a: 3) hypothesized that Plio-Pleistocene cooling was the major factors for the disappearance of endemic bee lineages form Hispaniola and the Greater Antilles.
The lack of closely related ancestors of A. poinari , in the Greater Antilles and Central America probably represents a manifestation of geographic extinction as described by Grimaldi (1992: 179), extinctions likely caused by drastic climatic changes as discussed by Poinar (1999 a: 3). Alternatively, the extinctions of these evolutionary intermediates may have been caused by bolide impact in the Yucatanian region at the end of the Cretaceous (Alvarez and Asaro 1990).
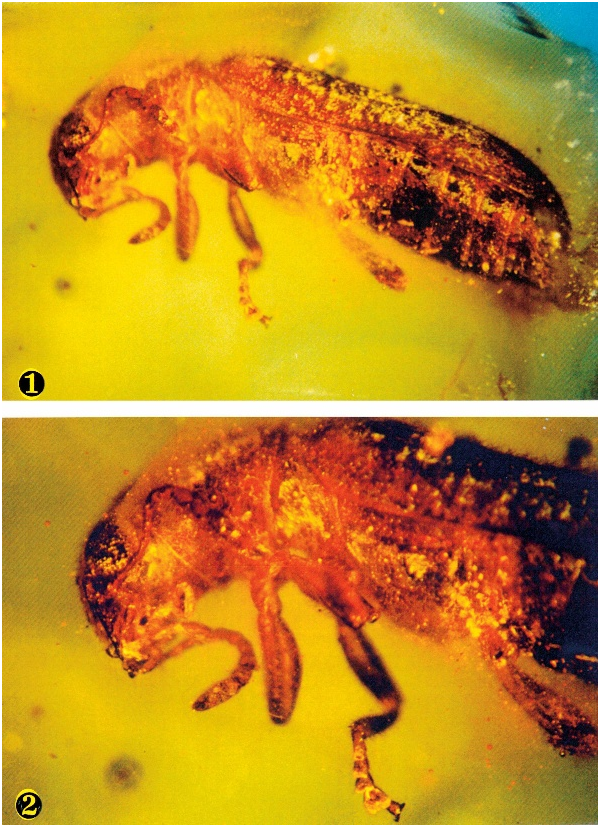
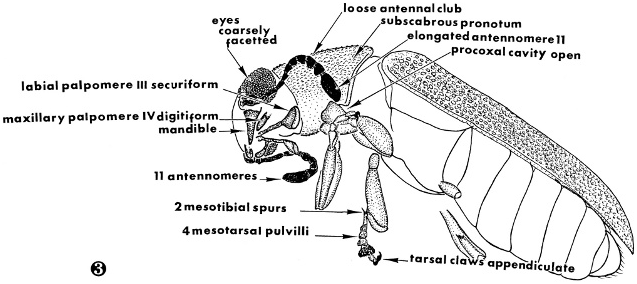
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |