
Aymaria, HUBER, 2000
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2000)254<0001:NWPSAP>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03ACD276-8FCC-FF74-FC8B-FA80437C3C5C |
|
treatment provided by |
Felipe |
|
scientific name |
Aymaria |
| status |
gen. nov. |
AYMARIA View in CoL View at ENA , NEW GENUS
TYPE SPECIES: Coryssocnemis conica Banks, 1902 .
ETYMOLOGY: The generic name honors the Aymara people, a vast group among the aboriginal people of South America, who by 1600 were reduced to a fourth of their preconquest numbers by slavery under the Spanish empire.
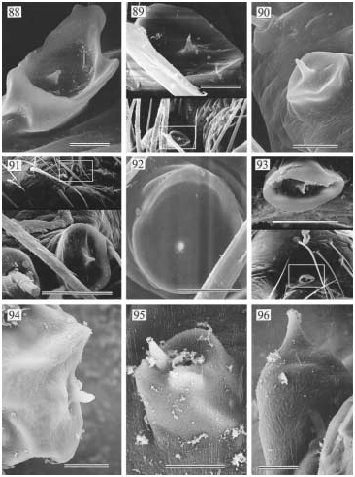
DIAGNOSIS: Medium-sized to large (total length 3 5 mm), usually dark, eight-eyed pholcids (troglobites pale and eyeless), with medium-long legs; distinguished from similar genera (e.g., Ixchela , n. gen., Priscula Si- mon) by the shape of the apophyses frontally on the male chelicerae overhanging the fangs (fig. 587; only in A. pakitza , n. sp., are they too small to overhang the fangs: fig. 601), the roughly circular indentation (pit) on the carapace, and the tent-shaped position of the pore plates in the female internal genitalia (figs. 592, 600, 605).
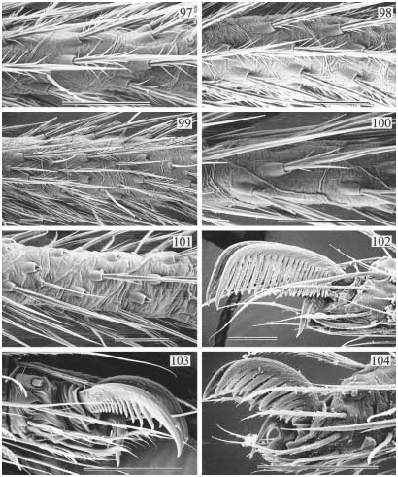
DESCRIPTION: Total length ~ 3 5 mm. Carapace with small to distinct round indentation (pit) behind ocular area; ocular area moderately elevated, with eight eyes, AME smallest; distance PME-ALE very large (~ 100 120% of PME diameter). Male clypeus unmodified. Male chelicerae with pair of distinctive apophyses distally, overhanging fangs (except in A. pakitza ); without stridulatory ridges laterally. Male sternum without humps. Male palpal coxa with retrolateral apophysis, femur with retrolateral apophysis proximally, ventral bulge distally (fig. 609); procursus very simple but with distinctive tip, bulb with apophysis distally on embolar division. Tarsal organ exposed (examined: A. conica : fig. 94). Legs relatively long (leg 1 about 6.5 11 × body length; tibia 1 l/d: 34 83), leg formula 1423 (legs 2 and 4 almost of same length); usually without spines, without curved and vertical hairs ( A. pakitza with curved hairs); retrolateral trichobothrium of tibia 1 very proximal (at ~ 5 6%); tarsus with 25 or more pseudosegments, which are very distinct distally. Opisthosoma high, either rounded, angular, or pointed posteriorly, with large dark spots dorsally. Male gonopore without epiandrous spigots (examined: A. conica ). ALS with only one piriform gland spigot each (examined: A. conica ), other spinnerets typical for family.
Sexual dimorphism slight. Epigynum consisting of anterior and large posterior plate (figs. 591, 604); pore plates in tentlike position (figs. 592, 600, 605). Some species with paired stridulatory apparatus dorsally between prosoma and opisthosoma (in females only).
MONOPHYLY: The species included share the unique position of the pore plates in the female internal genitalia; they further share many details in the shape of the pedipalp (the only interspecific differences occur on the procursus tip and the embolar division of the bulb), and are extremely similar in overall shape.
GENERIC RELATIONSHIPS: This genus shares several apomorphies with typical New World pholcids (exposed tarsal organ, male palpal coxa with retrolateral apophysis, large distance PME-ALE, male gonopore without epiandrous spigots, ALS with only one piriform gland spigot each), and seems particularly close to Ixchela , but this is only based on habitus, large size, and the simple, point- ed procursus. The pit in the prosoma is similar to that in the Old World Holocnemus group (and Artema and some Physocyclus ), but this is here interpreted as convergence.
SPECIFIC RELATIONSHIPS: The four nominal species from the Galápagos Islands have apparently indistinguishable genitalia, and are thus almost certainly a monophyletic group (two of these species may actually be synonyms; see Notes under A. conica below). They also share the stridulatory apparatus in the female, but this is also present in an undescribed species from Peru (Cajamarca, San Benito ), in MUSM .
DISTRIBUTION: Widely distributed from northern Argentina to Ecuador, and on the Galápagos Islands (map 4).
COMPOSITION: The genus as construed here includes seven nominal species. Of these, A. conica ; calilegua , n. sp.; pakitza n. sp.; and dasyops (Mello-Leitao) are treated below. A. insularis (Banks) , jarmila (Gertsch and Peck) , and floreana (Gertsch and Peck) are discussed under A. conica . I have seen two additional species, one from Peru (see above), and one from Ecuador, with much longer male cheliceral apophyses, but otherwise very similar to A. conica (deposited in CAS; poorly preserved).
Aymaria conica (Banks, 1902) , new combination Figures 94 View Figs , 103 View Figs , 581 View Figs 592
Coryssocnemis conica Banks, 1902: 56 ; pl. 1: figs. 1 3. Roth and Craig, 1970: 118. Baert and Maelfait, 1986: 107. Baert et al., 1989a: 45; 1989b: 16 17. Baert et al., 1991: 338.
Gertsch and Peck, 1992: 1186 1187, figs. 1 6, 12 14.
TYPES: Male lectotype, and a female prosoma only, from Hood Island (Isla Espanola), Galápagos Islands, Ecuador ; May 18, 1899 ( Hopkins Stanford Galápagos Expedition), in AMNH, examined .
DIAGNOSIS: Distinguished from A. calilegua , pakitza , and dasyops by the pointed end of the procursus (fig. 588); from A. pakitza also by the shape of the epigynum (fig. 591) and the larger apophysis on the male chelicerae (fig. 587); from A. insularis supposedly by the posteriorly pointed opisthosoma and the shorter legs ( A. insularis may in fact be a synonym of A. conica ; see Notes below); from A. jarmila and A. floreana by the presence of eyes.
MALE (lectotype): Carapace width 1.3; tibia 2: 4.8.
The following data are from a male from Santa Cruz Island: Total length 3.2, carapace width 1.5; leg 1: 27.2 (7.2+0.7+6.9+10.4 +2.0), tibia 2: 4.3, tibia 3: 3.3, tibia 4: 4.7; tibia 1 l/d: 41. Habitus and prosoma shape as in figs. 581, 584 586; distance PME-ALE about 120% of PME diameter. Carapace light brown with darker margins, ocular area and clypeus darker brown, sternum light brown. Chelicerae light brown with pair of dark brown apophyses overhanging fangs (fig. 587). Palps as in figs. 582 583, light brown; coxa with distinct retrolateral apophysis, femur with proximal protrusion and distal bulge, procursus with pointed end, with transparent membranous elements at tip (fig. 588), bulb with spinelike apophysis distally (fig. 589). Tarsal organ exposed (fig. 94). Legs light brown, with dark rings on femora (subdistally) and tibiae (proximally and subdistally); legs without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 6%; tarsus 1 with ~ 27 pseudosegments. Opisthosoma pale ochre, with many dark spots posteriorly and dorsally; gonopore without epiandrous spigots; ALS with only one piriform gland spigot each.
FEMALE: Tibia 1 in 13 females with rather pointed opisthosoma: 3.3 8.3 (!) (x¯ = 5.1). In general very similar to male. Epigynum usually very elevated (fig. 590), but this varies considerably (probably depending on amount of sperm in uterus externus); internal genitalia with pair of pore plates in tentlike position (fig. 592). Stridulation between pair of roundish humps posteriorly on carapace and pair of granulated plates frontodorsally on opisthosoma. The striated region frontally on epigynum (fig. 591; called rasping grate by Gertsch and Peck, 1992) has probably nothing to do with stridulation.
VARIATION: Tibia 1 in eight males: 4.9 10.7 (x¯ = 7.7). The opisthosoma apparently varies from rounded to pointed with all intermediates (see Notes below).
DISTRIBUTION: Apparently on most or all Galápagos Islands, as well as in Ecuador (mainland) and northern Peru (Piura) (map 4).
MATERIAL EXAMINED: ECUADOR, GAL- APAGOS ISLANDS: Española Island : types above ; Punta Cevallos, Jan. 5, 1983 (Y. Lubin), 13 1 juvenile in MCZ ; San Cristobal:
vic. El Junco (crater lake), ~ 700 m elev., Apr. 15 16, 1970 (R. Silberglied), 23 2♀ (4 vials) in MCZ ; same data, in lichens and mosses on Psidium guayaba , 13 in MCZ ; Santa Cruz: vic. Mirador (W of Media Luna), May 26, 1970 (R. Silberglied), 1♀ in MCZ ; pampas region, ~ 2 km W & N Media Luna , ~ 700 m elev., June 4, 1970 (R. Silberglied), 13 1 juvenile (2 vials) in MCZ ; above Media Luna, 1950 ft elev., Miceniazone , Apr. 17, 1981 (Y. Lubin), 1♀ in MCZ ; S. Plaza: 5 m elev., in dead fallen Opuntia sp. , Mar. 6 7, 1970 (R. Silberglied), 1♀ in MCZ ; Isla Rabida : May 12, 1981 (Y. Lubin), 1♀ 1 juvenile in MCZ ; Isla Santiago: E Side: Los Guayabitos , from lava tube, Feb. 27, 1983 (collector not given), 13 in MCZ ; SE Cumpleanos (tortoise nesting zone C), Mar. 11, 1983 (C. Marquez), 1♀ in MCZ ; Santa Fé : S Coast, Jan. 28, 1983 (Y. Lubin), 1♀ in MCZ ; ECUADOR (mainland): Puná Island, Jan. 23 26, 1941 (D.L.F.), 23 in CAS ; Guabillo , 20 km W Arenillas, Oct. 29, 1942 (no collector given), 1♀ assigned ten- tatively, in CAS ; PERU: Piura: Quebrada Mogollon (4°32'S, 81°04'W), June 11, 1939 (D.L.F. & H.E. F.), 23 1♀ in MCZ GoogleMaps ; 12 km SW Fernández (~ 4°12'S, 80°51'W), Jan. 1, 1939 (D.L.F. & H.E. F.), 13 3♀ 2 juveniles GoogleMaps
in MCZ; Canchaque (5°24'S, 79°36'W), ~ 800 m elev., Mar. 24, 1989 (D. Silva) 33 3♀ in MUSM GoogleMaps .
NOTES: The main point in the present context is that the four Galápagos species previously included in Coryssocnemis Simon are not closely related to the type species C. callaica Simon , but are part of a distinct group of species (herein considered a genus) widely distributed in western South America. With the material examined, I am reluctant to make statements concerning the number and diagnostic characteristics of the Galápagos species involved. At least two species occur on the islands, one epigean with eight well-developed eyes, and one completely eyeless cavernicole species. The genitalia are surprisingly similar (in fact, I have been unable to detect any genitalic differences, see below), but the complete absence of eyes in one group and the absence of intermediate forms strongly suggests that the two are reproductively isolated. The more confusing problem is that each group is currently again separated into two nominal species: the epigean A. conica (Banks) and A. insularis (Banks) , and the blind A. jarmila (Gertsch and Peck) and A. floreana (Gertsch and Peck) . I suspect that one name in each group is a synonym, but more material should be studied and rigorously compared to allow a convincing solution. The following hints may be useful for future studies at the species level.
Banks (1902) described the two epigean species, and noted that in insularis the legs are much stouter and longer than in [ conica ], and the abdomen is not prolonged above into a pointed cone, but broadly rounded behind, although it projects considerably over the spinnerets. Gertsch and Peck (1992) (in a paper that is full of contradictions, typing and other errors) added figures of the female genitalia supposedly showing diagnostic differences. All three proposed differences seem dubious: leg length and stoutness vary widely, as does the shape of the opisthosoma, and I have been unable to understand how Gertsch and Peck (1992) managed to see any epigynal differences. In fact, KOH preparations of the genitalia of the insularis holotype and another female identified by Gertsch as insularis showed no differences to prepara- tions made from typical (i.e., pointed abdomen) conica females. The two supposed species are not geographically separated (they share several islands, and at least once, even the same cave).
The two cavernicole species were said to differ only in the female genitalia: in floreana the frontal rasping grate was said to be undivided, but divided in jarmila , and internal differences were figured by Gertsch and Peck (1992). Again, I have been unable to see these differences. KOH preparations of the genitalia of the holotypes of jarmila and floreana revealed no differences; both appear similar if not identical to the epigean species, and show little resemblance to the figures in Gertsch and Peck (1992). Finally, in one female from Floreana Island I examined (i.e., supposedly floreana ), the rasping grate was clearly divided into two parts (i.e., as in jarmila ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.