
Gymnothorax elaineheemstrae Sithole, Smith & Gouws, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4767.1.7 |
|
publication LSID |
urn:lsid:zoobank.org:pub:7851D34D-A3ED-4F1C-A5AD-5AF43D35E0AF |
|
DOI |
https://doi.org/10.5281/zenodo.3798898 |
|
persistent identifier |
https://treatment.plazi.org/id/9B538791-FC52-FF8C-E2D7-340E5CCAA680 |
|
treatment provided by |
Carolina |
|
scientific name |
Gymnothorax elaineheemstrae Sithole, Smith & Gouws |
| status |
n. sp. |
Gymnothorax elaineheemstrae Sithole, Smith & Gouws n. sp.
Common name: Marbled leopard moray
Figs. 3 View FIGURE 3 , 5 View FIGURE 5 ; Table 9 View TABLE 9
Lycodontis undulatus (non Lacepède 1803): Smith 1949: 398 (Fig. 1129, Pl. 100); Smith 1953: 398 (Fig. 1129, Pl. 100); Smith 1962: 439 (Fig. E, Pl. 57); Castle & McCosker 1986: 173 (Fig. 41.24, Pl. 8).
Gymnothorax cf. undulatus: King & Fraser 2014: 34 , 35 (Fig. unnumbered)
Holotype: SAIAB 86307 (624 mm TL), South Africa, KwaZulu-Natal, Cape Vidal , 28°9.8'S, 32°34.1'E, 6 Jan. 2009, coll. A. Paterson, G. Musson and A. Goetz. GoogleMaps
Paratypes: SAIAB 206655 (383 mm TL), same as holotype; CSIRO H 8399-01 (304 mm TL), South Africa, KwaZulu-Natal, Pennington, tide pools in front of the golf course; 30°24'S, 30°42'E, 29 Jul. 1992; AMS I.48960- 001 (665 mm TL), South Africa, Eastern Cape, Coffee Bay, 32°00'S, 29°12'E, 20 Jan. 1976; USNM 330982 (454 mm TL), South Africa, Natal, Aug. 1992.
Non-type material: SAIAB 46644 (2: 285–329 mm TL), South Africa, KwaZulu-Natal, Mbibi, 27°30'S, 32°42'E, 12 Dec. 1988; SAIAB 4990 (2: 325–343 mm TL) South Africa, Eastern Cape, Mtentwana Point, 31°06'S, 30°12'E, 15 Aug. 1958; SAIAB 83082 (710 mm TL) South Africa, KwaZulu-Natal, Scottburgh 30°00'S, 31°00'E, 05 Jun. 2007; SAIAB 44497 (485 mm TL) South Africa, Eastern Cape, Algoa Bay, 34°00'S, 25°42'E, 01 Jan. 1967; SAIAB 39939 (644 mm TL) South Africa, Eastern Cape, Transkei Presley Bay, 31°54'S, 29°18'E, 01 Sep. 1974; SA- IAB 39959 View Materials (667 mm TL) South Africa, Eastern Cape, Hole-In-Hall, 32°00'S, 29°12'E, 27 Jan. 1975; SAIAB 83077 (2: 171–267 mm TL) South Africa, KwaZulu-Natal, Scottburgh, 30°17'S, 30°46'E and SAIAB 60166 (9: 85–349 mm TL), South Africa, KwaZulu-Natal, Scottburgh tide pool, 30°18'S 30°48'E, 18 Apr. 1999.
Diagnosis. A moray eel with dorsal-fin origin anterior to gill opening, anal-fin origin just behind anus, with mottled and faintly reticulated color pattern on a dark brown background; three long intermaxillary canines in a median row; vomerine teeth small, in a single row; maxillary teeth biserial; vertebral count: 3–5 predorsal, 54–58 preanal, and 134–136 in total.
Description. Proportions as % TL: preanal 41.1–50.8, predorsal 8.1–11.5, head length 11.3–15.0, depth at gill opening 4.2–8.4, depth at anus 3.7–6.5. As % head length: snout 15.2–18.5, eye diameter 10.0–11.8 and upper jaw 37.0–47.9. Vertebral: predorsal 3–5 (3), preanal 54–58 (56), total 134–136 (135).
Body elongate, depth at gill opening 4.2–8.4 and 3.7–6.5 at anus in % TL; anus anterior to mid-body, preanal length 41.1–50.8 in % TL; head and trunk together shorter than the tail. Dorsal-fin origin anterior to gill opening, predorsal length 8.1–11.5 in % TL; anal-fin origin immediately behind anus; dorsal and anal fins continuous. Head long 11.3–15.0 in % TL, blunt anterior, concave above eye and broad behind the eye. Snout short, 15.2– 18.5 in % HL; upper jaw slightly longer than the lower jaw; anterior nostril tubular on each side of snout tip, posterior nostril round and slightly anterior to the eye.
Three supraorbital pores, first pore situated just above the upper lip near tip of snout, second above the first pore and adjacent to the anterior nostril and one between anterior and posterior nostrils. Two branchial pores above and anterior to gill opening. Four infraorbital pores along the upper jaw, first between anterior nostril and anterior margin of the eye, second below anterior margin of the eye, third below the middle of the eye and the fourth below posterior edge of the eye. Five mandibular pores, all anterior to the rictus. No lateral-line pores posterior to the branchial ones. Gill opening slit-like and slightly below mid-body.
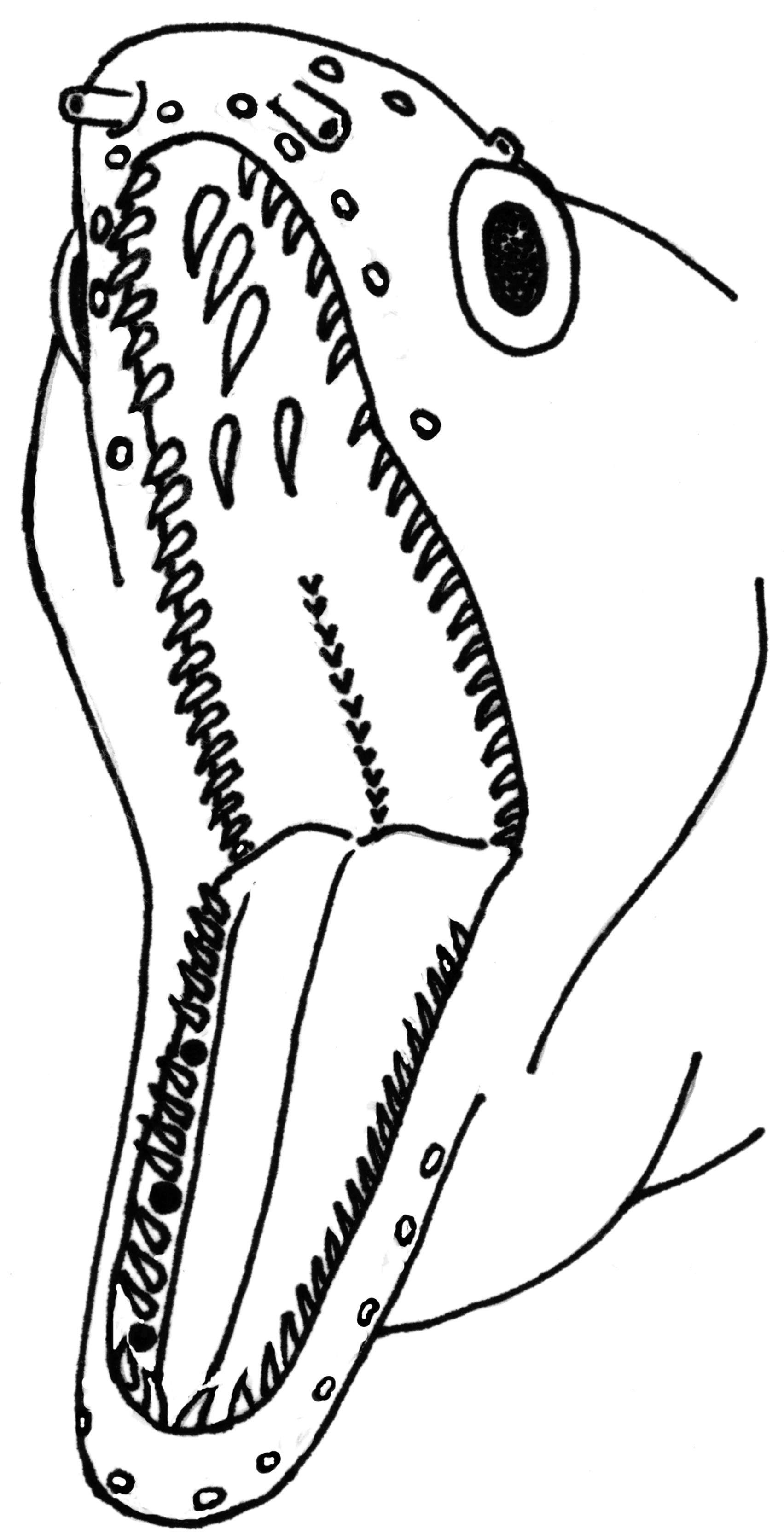
Teeth slender and sharply pointed ( Fig. 3 View FIGURE 3 ); peripheral inter-maxillary teeth in single row of 6–9 (6) on each side; no intermediate inter-maxillary teeth; median inter-maxillary 2–3 (3), uniserial extremely long and widely spaced; vomerine teeth uniserial and small; inner maxillary teeth 1–4 (1), longer than outer maxillary teeth, anterior ones the largest, decreasing in size posteriorly; outer maxillary 12–16 (14) small teeth, closely spaced.
Colour of fresh specimen and in preservation ( Fig. 5 View FIGURE 5 ). Head and body dark brown with mottled and faintly reticulated pattern; mottled coloration on the head extends inside the lower jaw; snout and ventral portion of the lower jaw to the beginning of anal fin paler compared to the rest of the body; dorsal- and anal-fins with dark brown margins.
Distribution. Specimens of this species were collected in the Western Indian Ocean along the coast of South Africa from KwaZulu-Natal to the Eastern Cape, known to a depth of ~ 40 m in rock, intertidal and tide pools and sand bottoms.
Etymology. The specific name, elaineheemstrae , is in honour of Mrs Elaine Heemstra (NRF-SAIAB Honorary Research Associate) for her tremendous support in mentoring the first author in fish taxonomy.
Comparisons with other species. The dorsal-fin origin anterior to gill opening and anal-fin origin just after anus places this species in the genus Gymnothorax as currently recognized. Gymnothorax elaineheemstrae can be differentiated from other species within the genus in the Western Indian Ocean by its colour pattern and vertebral count. Gymnothorax elaineheemstrae has a mottled and faintly reticulated pattern ( Fig. 5 View FIGURE 5 ), whereas G. undulatus has irregular rectangular blotches separated by large reticulations ( Fig. 4 View FIGURE 4 ). Gymnothorax elaineheemstrae has a similar teeth arrangement to G. chilospilus Bleeker 1864 , an Indo-Pacific species which G. undulatus has been confused with. However, G. chilospilus differs from G. elaineheemstrae in having fewer preanal (47–52 vs 54–58) and total vertebrae (120–129 vs 134–136); more peripheral intermaxillary teeth (6–8 vs 4–6) and clearly visible white markings on head (vs no white markings on head). The mottled pattern in G. elaineheemstrae is similar to G. flavimarginatus ( Rüppell 1830) , but G. elaineheemstrae has a faintly reticulated pattern and lacks the black blotch at the gill opening and the pale margin around posterior tail. Gymnothorax elaineheemstrae also differs from another closely similar species G. pictus ( Ahl 1789) by the dorsal-fin origin which is anterior to gill opening (vs exactly above gill opening in G. pictus ); uniserial teeth on vomer (vs. biserial); 3–5 (vs 7–11) predorsal and 54–58 (vs 56–62) preanal vertebrae.
Remarks. The species retained the common name Marbled leopard moray given by King & Fraser (2014).
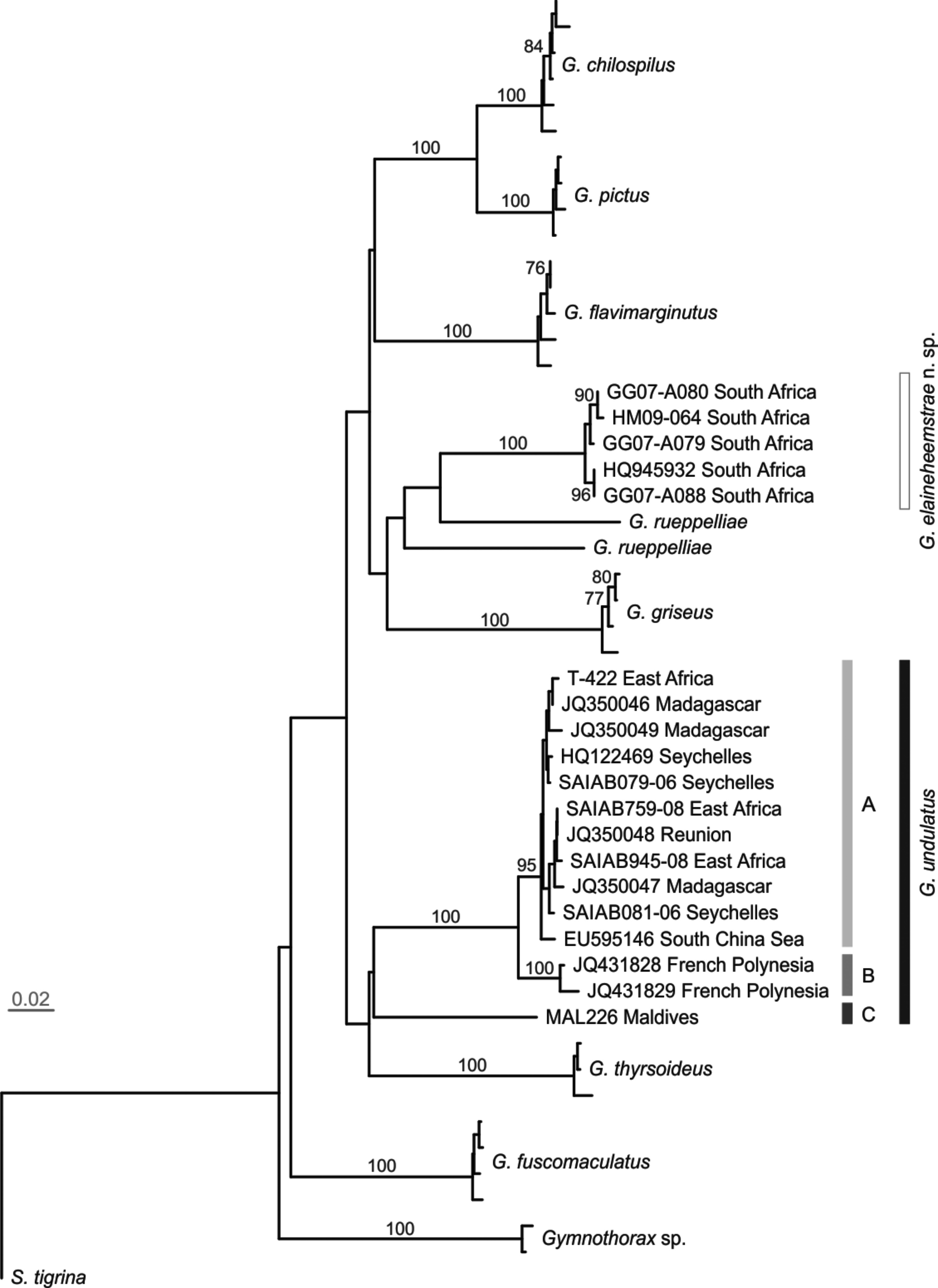
Genetics. The COI analysis gave an aligned data set of 411 base pairs (bp) in length for 50 sequences. The neighbour-joining tree ( Fig. 6 View FIGURE 6 ) based on COI sequences revealed two distinct clades among specimens formerlyregarded as G. undulatus . Gymnothorax undulatus was made up of three lineages (Clades A, B and C) and was geographically widespread, comprising samples from the Pacific Ocean and several WIO regions. These lineages represented regions that were not distinct in the morphological analysis. Specimens from all these geographic regions had the typical G. undulatus color pattern, i.e. dark brown body with rectangular spots separated by pale reticulum. The second clade comprised Gymnothorax elaineheemstrae and was nested within the morphologically-distinct banded moray G. rueppelliae ( McClelland 1844) . Gymnothorax elainheemstrae is restricted to South Africa and comprises of specimens that are mottled and less reticulated. The most appropriate nucleotide substitution model for the COI data was determined to be a GTR model ( Rodríguez et al. 1990), with proportion of invariable sites (I = 0.638), base frequencies of A = 0.284, C = 0.290, G = 0.157 and T = 0.269, and a rate matrix of R [A↔C] = 1.247, R [A↔ G] = 19.982, R [A↔T] = 3.994, R [C↔G] = 1.790, R [C↔T] = 18.676 and R [G↔T] = 1.000. The ML tree constructed with a loglikelihood (lnL) of -2829.731 (not shown) indicated the genetic distinction of G. undulatus and G. elaineheemstrae , similar to neighbour joining tree. Enforcing the topology to reflect the monophyly of the former G. undulatus (i.e., G. undulatus + G. elaineheemstrae as sister taxa, maintaining all internal relationships) resulted in a log-likelihood of -2840.998; this topology was significantly less likely (∆lnL = 11.266, P <0.05) than the optimal topology. The average pairwise K2P interspecific genetic distances between examined species of Gymnothorax based on COI sequences ranged from 0.073 to 0.267 ( Table 10 View TABLE 10 ). The lowest genetic distance (0.073) was between the paintspotted moray G. pictus and lipspot moray G. chilospilus , and this was the only genetic distance <0.10. The highest (0.267) was between the geometric moray G. griseus and brown spotted moray G. fuscomaculatus . The genetic distance (K2P) between G. undulatus and G. elaineheemstrae was high at 0.201.
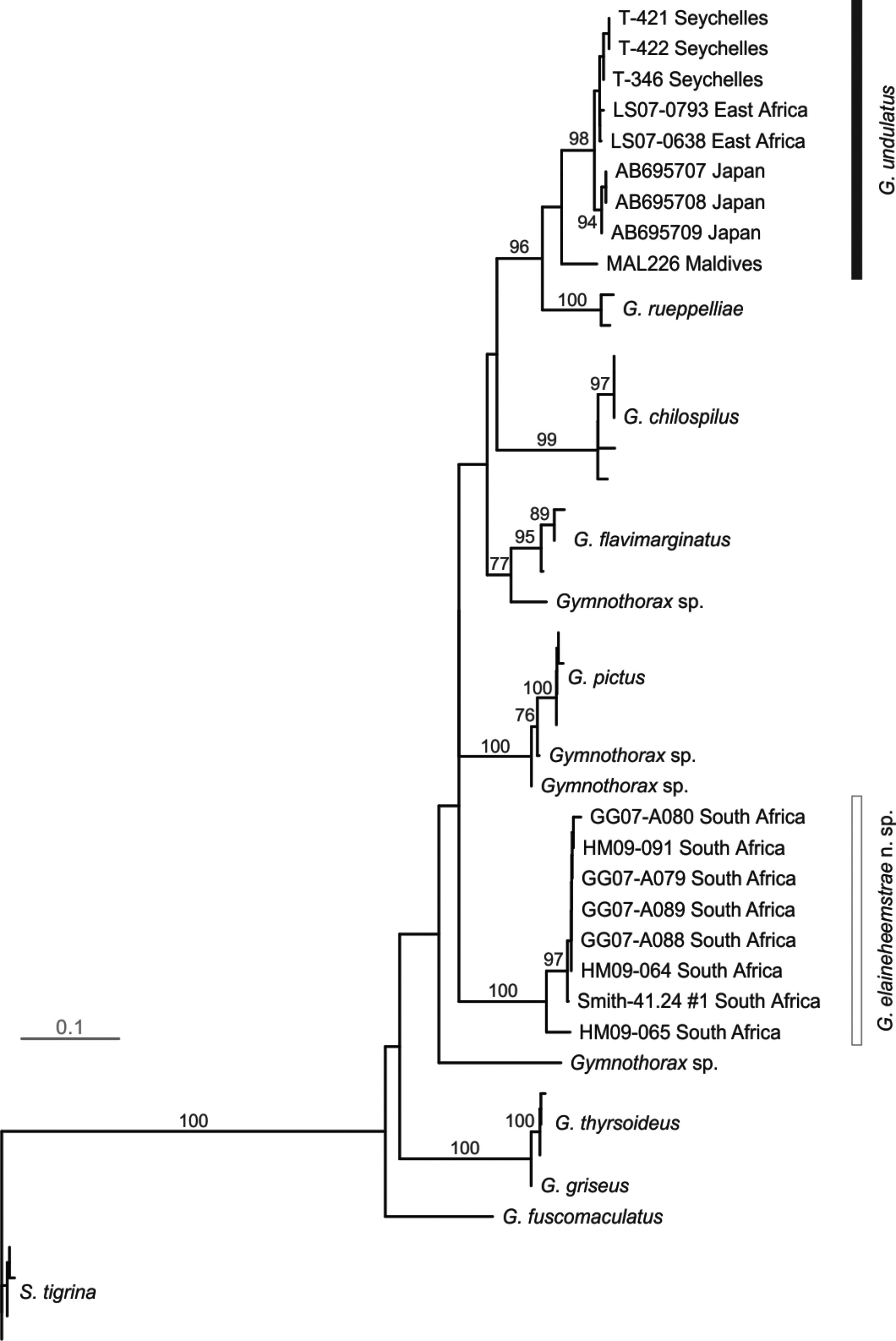
The 16S rRNA analysis involved an alignment of 404 nucleotides for 44 sequences, including three sequences of G. undulatus from Japan (Pacific Ocean). The optimal model for this dataset included a model with base frequencies of A = 0.339, C = 0.260, G = 0.196 and T = 0.205, a rate matrix of R [A↔C] = 1.277, R [A↔G] = R [C↔T] = 5.725 and R [A↔ T] = R [C↔G] = R [G↔T] = 1.000, a proportion of invariable sites (I = 0.367) and a gamma distribution of rate variation (α = 0.477). The NJ tree ( Fig. 7 View FIGURE 7 ) recovered a clade of G. undulatus that was widespread, containing samples from the Archipelago, East Africa, Seychelles and Japan, and sister to G. rueppelliae . Gymnothorax elaineheemstrae was distinct and phylogenetically divergent from the G. undulatus clade and was well-supported with 100% bootstrap. The ML tree (lnL = -2490.389; not shown) also revealed G. undulatus and G. elaineheemstrae to form distinct and phylogenetically well-separated clades. As with the COI data, enforcing the topologies to reflect the monophyly of the former G. undulatus (with G. undulatus placed as a sister taxon to G. elaineheemstrae ) presented a significantly worse fit to the data (lnL = -2512.244, ∆lnL = 21.855, P <0.05) than the optimal topology. The average model-corrected pairwise sequence divergences between examined species based on 16S rRNA sequences are given in Table 11. The highest genetic distance (0.408) was between the greyface moray G. thyrsoideus and the banded moray G. rueppelliae , and the lowest value (0.003) was between G. griseus and G. thyrsoideus , possibly indicating the misidentification of the former (see Fig. 7 View FIGURE 7 ). The next lowest value (0.190) was between G. chilospilus and G. flavimarginatus . The genetic distance from the comparison between G. elaineheemstrae and the G. undulatus clades was 0.234. Average divergences of 0.021 to 0.081 separated potential Archipelago, widespread and Japanese (Pacific Ocean) subclades and lineages within the G. undulatus clade ( Fig. 7 View FIGURE 7 ).


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gymnothorax elaineheemstrae Sithole, Smith & Gouws
| Sithole, Yonela, Smith, David G., Mwale, Monica & Gouws, Gavin 2020 |
Gymnothorax cf. undulatus: King & Fraser 2014: 34
| King, D. & Fraser, V. 2014: 34 |
Lycodontis undulatus
| Castle, P. H. J. & McCosker, J. E. 1986: 173 |
| Smith, J. L. B. 1962: 439 |
| Smith, J. & LB 1953: 398 |
| Smith, J. L. B. 1949: 398 |
| Lycodontis undulatus (non Lacepède 1803 |