
Lasionycta Aurivillius, 1892
|
publication ID |
https://doi.org/10.3897/zookeys.30.308 |
|
publication LSID |
lsid:zoobank.org:pub:C26E1A82-0DD4-48EF-865C-9D8AA788B739 |
|
DOI |
https://doi.org/10.5281/zenodo.3790271 |
|
persistent identifier |
https://treatment.plazi.org/id/75513F41-7B72-FFDF-FF02-ED19930CFADF |
|
treatment provided by |
Plazi |
|
scientific name |
Lasionycta Aurivillius |
| status |
|
Lasionycta Aurivillius View in CoL
Lasionycta Aurivillius, 1892b: 285 View in CoL . Type species: Phlogophora skraelingia Herrich- Schäffer by original designation.
Lasiestra Hampson, 1905: 47 View in CoL . Type species: Dianthoecia phoca Möschler by original designation.
Pseudanarta Kozhanchikov, 1947: 18 View in CoL . Type species: Anarta bohemani Staudinger by original designation. Preoccupied by Pseudanarta Grote, 1878 View in CoL .
Anartomima Boursin, 1952: 55 View in CoL . Replacement name for Pseudanarta Kozhanchikov. View in CoL
Diagnosis. Lasionycta is placed in Hadeninae based on trifine hindwing venation (vein M2 vestigial), hairy eyes, and larva with a dorsoventrally flattened spinneret with the opening shaped as a narrow transverse slit with an apical fringe. Within Hadeninae , Lasionycta is classified in Eriopygini Fibiger and Lafontaine based on an undivided larval hypopharynx, lack of an inner tooth on the larval mandible, and features of the larval spinneret, including elongated shape and apical fringe confined to the end of groove ( Fibiger and Lafontaine 2005).
Within Eriopygini, Lasionycta is characterized by the male genitalia, which have the following features: 1) an uncus with a nearly cylindrical base with a pointed or dorso-ventrally flattened tip; 2) a strap-like valve bearing a C-shaped clasper, an elongate digitus oriented 30–90° to the axis, a small to moderately large cucullus with a simple or compound corona, and a membranous flap on the mesial surface distal to the sacculus (post-saccular flap); 3) an elongate coiled or spiraled vesica bearing zero to six pointed subbasal cornuti with broad bases and a distal field of minute cornuti or one or two rows of spine-like cornuti oriented perpendicular to the vesica axis. The distal cornuti field is adjacent to a membranous wrinkled band running the length of the vesica.
Description. Head – Male antenna beadlike, biserrate, or bipectinate (1.1 – 5.4× as wide as central shaft) and fasciculate. Female antenna filiform and ciliate. Scape with a tuft of longer scales at dorsal base of antenna. Eye hairy, round to ellipsoid. Head covered in hair-like scales. Thorax – Vestiture dense hair-like scales or mixture of hair-like and weakly spatulate scales. Foretibia with epiphysis. Tarsal segments with three rows of spine-like setae. Wings – Ground color light to dark gray or gray brown. Wings of most species with typical noctuine lines and spots present (indistinct in several species). Hindwing nearly uniform gray or lighter gray, white, or yellow with variable dark suffusion and contrasting darker discal spot, postmedial line, and marginal band. Male genitalia – Uncus curved, slightly laterally compressed with hook-like apex or with distal portion dorsoventrally flattened with ovoid or spatulate apex. Valve elongate, strap-like, 4.5–8.0× as long as wide. Sacculus rounded, relatively weak, with costal lobe extending below or above costal margin of valve. Valve bearing a membranous flap on ventromesial surface distal to sacculus ("postsaccular flap") (lacking in L. secedens species-group). Distal portion of valve weakly to moderately constricted below cucullus (neck). Cucullus 0.65–1.50× as wide as valve, with corona of 1–4 rows of stout claw-like setae. Clasper cylindrical and curved posterodorsad. Digitus moderately sclerotized, oriented 30–45° ventrad, finger-like with variable width base, or more heavily sclerotized, oriented 90° ventrad, with a basal flange or prong ( L. subdita species-group). Aedeagus cylindrical, tubular, or with single thorn-like or multiple-tipped raised apical projection on right; vesica 1.5–3.0× as long as aedeagus, curved and twisted to nearly straight beyond a subbasal bend or coil, distal part extending right and ventral to distal aedeagus, with zero to six basal cornuti (variable between and within species) comprised of a central spike from a smooth to crenulate button-like base; vesica with a wrinkled lightly sclerotized band along entire length, distal vesica with adjacent field of multiple setae as a band of in- numerable velvety setae or one or more rows of stouter setae oriented perpendicular to vesica axis. Female genitalia – Ovipositor lobe membranous and pad-like covered with hair-like setae or stout and conical covered with short stout setae. Abdominal segment VIII 0.3–1.2× as long as wide, short in species with pad-like ovipositors and elongate in species with conical ovipositors, covered with variable number of short setae, greatest ventrally and along posterior margin. Ductus bursae moderately sclerotized, cylindrical to dorsoventrally flattened, distal ventral aspect turned onto adjacent dorsal membranous bursa as a flange-like supporting structure; some species groups with posterior end heavily sclerotized and expanded to left ( L. secedens species-group) or anterior end expanded to right ( L. subdita species-group). Corpus bursae membranous, spherical or ovoid, with 3 elongate signa separated by 90°, one on each surface except side with appendix bursae (1 ventral signum in L. secedens species-group), with very weak to prominent mesial constriction, most pronounced on dorsal side near appendix bursae origin. Appendix bursae membranous or lightly sclerotized, arising from left side ( L. skraelingia species-group) or dorsum of posterior corpus bursae and extending anteriorly to left, or posterodorsad from corpus bursae; shape of appendix weakly to strongly coiled 570° ( L. phaea and L. secedens species-groups, respectively) or straight with subapical 90° bend. Ductus seminalis at distal appendix bursae (posterior end in most species, anterior end in L. skraelingia species-group).
Early stages. Larvae are known for L. skraelingia (Herrich-Schäffer) , L. secedens (Walker) , L. leucocycla (Staudinger) , L. staudingeri (Aurivillius) , and L. perplexa (Smith) ( Godfrey 1972; Ahola and Silvonen 2008). Hypopharynx undivided, mandible lacking a tooth on inner surface, spinneret long and narrow, and fringe at spinneret apex confined to a small area at end of dorsal groove ( Fibiger and Lafontaine 2005).
Remarks. The Lasionycta species in this revision are included in Anartomima Boursin , Lasiestra Hampson , and Lasionycta in the Moths of North America checklist ( Franclemont and Todd 1983). Lasiestra ( type species: Dianthoecia phoca Möschler ) has since been synonymized with Lasionycta ( Lafontaine et al. 1986) . Lasionycta secedens was first treated as a Lasionycta in Europe ( Hacker et al. 2002). It resembles L. skraelingia and L. phaea (Hampson) and like them the vesica is long; superficially it differs mainly in hindwing color. Its generic placement was scrutinized for this work because both sexes of L. secedens differ from other Lasionycta by several seemingly unique anatomic features. Characters intermediate between L. secedens and other Lasionycta species were found in the L. phaea and L. mutilata species-groups. Also, CO1 gene sequence distance analysis nests L. secedens among other Lasionycta speciesgroups, usually closest to the L. phaea species-group. This supports its inclusion in Lasionycta , and it is herein treated as the sole member of the L. secedens species-group. A check list of the North American Lasionycta species is presented in Table 2.
Species included in Lasionycta in recent European literature are more diverse in appearance, morphology, and biology than in North America. Hacker et al. (2002) list 18 Palaearctic species in the genus. Four of these ( L. skraelingia , L. leucocycla , L. staudingeri , and L. secedens ) are Holarctic and are included in the current revision. Two other Asian species, L. buraetica Kononenko and L. corax Kononenko , are related to Nearctic L. phaea , as is L. alpicola Lafontaine & Kononenko ( Lafontaine and Kononenko 1988) which was omitted from this list. Four other species, proxima , calberlai (Staudinger), imbecilla (Fabricius) , and impar (Staudinger), occur in Europe and were described and illustrated by Hacker et al. (2002). Th ey have no close relatives in North America and were therefore evaluated to determine their relationship to other Lasionycta .
Proxima, the type species of Lasionhada Berio, 1981 , has genitalia (Figs 190 and 245) similar to other Lasionycta . Th e male genital capsule resembles closely those of the Lasionycta leucocycla species-group in all characters including the presence of the post-saccular flap. However, the vesica differs from those of Lasionycta species in this revision in that the elongate band of distal setae are positioned on the membranous band, not at its margin. Th e female genitalia are similar to those of the L. leucocycla species-group but lack signa on the corpus bursae and have long symmetrically pointed ovipositor lobes, characters not found elsewhere in Lasionycta . Th e CO1 sequence of proxima suggests that it is an outlier from other Lasionycta species, usually clustering adjacent to them but occasionally grouping with related genera like Tricholita Grote ( Fig. 247 View Figure 247 ). It also differs biologically from other Lasionycta in that it occurs in dry temperate areas as low as sea level, whereas other Lasionycta species prefer conifer forests or tundra. For these reasons we transfer proxima back to Lasionhada as Lasionhada proxima (Hübner) , comb. rev. Th e morphologic and DNA data suggest that Lasionhada is the sister group to Lasionycta .
Calberlai is the type species of Clemathada Beck, 1991 and was moved from Lasionycta back to this genus by Fibiger and Lafontaine (2005). Th ey noted that it belongs in Hadenini, not Eriopygini, based on larva characters including a divided hypopharynx.
Imbecilla is the type species of Eriopygodes Hampson, 1905 and was first associated with Lasionycta by Hacker et al. (2002). It was examined and differs significantly from Lasionycta in several respects, including presence of a stout broadly ovate clasper, a vestigial digitus, and no postsaccular flap of the male valve (Fig. 191). It does not group with other Lasionycta on CO1 sequence analysis, instead clustering with Lacinipolia lorea (Guenée) , which it also resembles superficially. Lacinipolia McDunnough remains a large heterogeneous genus in need of revision. Imbecilla is herein transferred back to Eriopygodes as E. imbecilla (Fabricius) , comb. rev.
Impar was not available for examination for this revision but is illustrated by Hack- er et al. (2002). Its habitus and vesica are somewhat atypical for Lasionycta although the genitalia are similar. We retain it in Lasionycta pending further review of the Eurasian Eriopygini.
The other eight species in the Hacker list ( L. hospita Bang-Haas , L. melanographa Varga , L. montanoides Poole, L. orientalis (Alpharéky) , L. decreta (Püngeler) , L. draudti (F. Wagner) , L. hampsoni Varga , and L. levicula (Püngeler)) are Asian and were not otherwise evaluated by Hacker et al. (2002) or by us in the current work. Only the first four of these species are included in Lasionycta by Poole (1989).
Four North American species included in Lasionycta in recent checklists (Franclemont and Todd 1982; Poole 1989) ( L. insolens (Grote) , L. arietis (Grote) , L. wy- atti (Barnes & Benjamin), and L. ochracea (Smith) and the recently described L. sala Troubridge & Mustelin ) occur on Pacific Ocean sand beaches. Th eir larvae and adults differ structurally from other Lasionycta . Godfrey (1972) grouped L. arietis and L. wyatti separately from other Lasionycta ( L. perplexa and an identified species of “ Lasiestra ” from Mt Rainier) in a comprehensive review of North American Hadenine larvae, recognizing that they formed a “very unique group.” These beach species cluster on CO1 distance analysis, consistently nesting among Lacinipolia species and removed from other Lasionycta . Th ese data indicate that they are not closely related to Lasionycta . A new genus, Psammopolia Crabo & Lafontaine , is described for them in this paper.
Many Lasionycta are alpine and have virtually no prospect for gene exchange between populations in different mountain ranges. It is not surprising that many populations differ slightly in appearance from those from nearby ranges. This revision required many decisions on whether to treat various levels of dissimilar disjunct populations as species, subspecies, or variation. After discovering convincing evidence that several L. leucocycla species-group taxa previously considered subspecies of L. leucocycla were species (e.g., L. poca (Barnes & Benjamin) and L. flanda (Smith)) greater weight was given to superficial characters of pattern and color, taking care not to over interpret variability in species like L. uniformis (Smith) . Proposed changes were tested against DNA data to formulate the most congruous final classification. Nonetheless, we usually gave greater weight to morphology and habitus than DNA and recognize several species without any DNA sequence differences, especially where the ranges were found to abut or overlap without any intermediate forms in the areas of sympatry. Taxa were treated as subspecies when there were significant differences in appearance between large regional populations with only slight overlap in characters, especially in species with multiple CO1 haplotypes in which barcode differences were considered to be less reliable than in groups with less variable DNA. We resisted the temptation to name small populations (e.g., L. promulsa (Morrison) from the Uinta and Wasatch Mountains), except when highly disjunct ( L. uniformis shasta ssp. n. in California and L. uniformis handfieldi ssp. n. in eastern Canada).
Barcode results. A total of 310 North American Lasionycta specimens were submitted for CO1 barcode sequencing. Of these, 208 produced sequences of 600 or more base pairs and 168 were complete (658 bp). No barcode results are available for four species ( L. carolynae , L. illima , L. macleani , L. mono ) and only incomplete sequences are available for eight more ( L. coracina , L. dolosa , L. flanda , L. lagganata , L. phaea , L. sasquatch , L. sierra , L. subfumosa ) (all barcode data is presented in the Appendix). The sample set is skewed by the availability of fresh material from the Pacific Northwest, Alberta, and Churchill, Manitoba and a relative paucity of suitable material from most other parts of North America. Most samples are from sub-groups in which variability suggested the possibility of several similar species. Lasionycta uniformis , L. subfuscula , L. perplexa , and L. leucocycla are particularly well sampled.
Twenty of the 39 species (51.3 %) with complete or partial sequences had a single CO1 haplotype. Five of these are only known from a single sequence, so the true haplotype diversity of these species is unknown. Eleven species (28.2 %) had two haplotypes, and two species had three (5.1 %). Th ere were no species with four or five haplotypes, but six species (15.4 %) had six to twelve haplotypes ( L. leucocycla , L. perplexa , L. promulsa , L. subfuscula , L. taigata , L. uniformis ). Eight of 39 species (20.5 %) shared haplotypes ( L. skraelingia and L. taigata , L. anthracina and L. flanda , L. poca and L. sasquatch , and L. coracina and L. leucocycla ). Th e haplotypes of 8 species differed by only one or two base pairs ( L. frigida and L. leucocycla , L. phoca and L. uniformis , L. pulverea and L. sierra , L. staudingeri and L. subfumosa ). The neighborjoining trees ( Figs 247 View Figure 247 and 248 View Figure 248 ) demonstrate that seven species are placed adjacent to or nested amongst multiple similar haplotypes of two polytypic species ( L. anthracina , L. flanda , L. frigida , and L. coracina with L. leucocycla ; L. phoca , L. gelida , and L. discolor with L. uniformis ). Therefore, 17 Lasionycta species (43.6 %) have identical or similar barcodes demonstrating a high frequency of non-discrete haplotype clusters despite the relatively small number of highly polytypic species. Additional sampling of poorly represented species is likely to show an increase in polytypism and haplotype sharing.
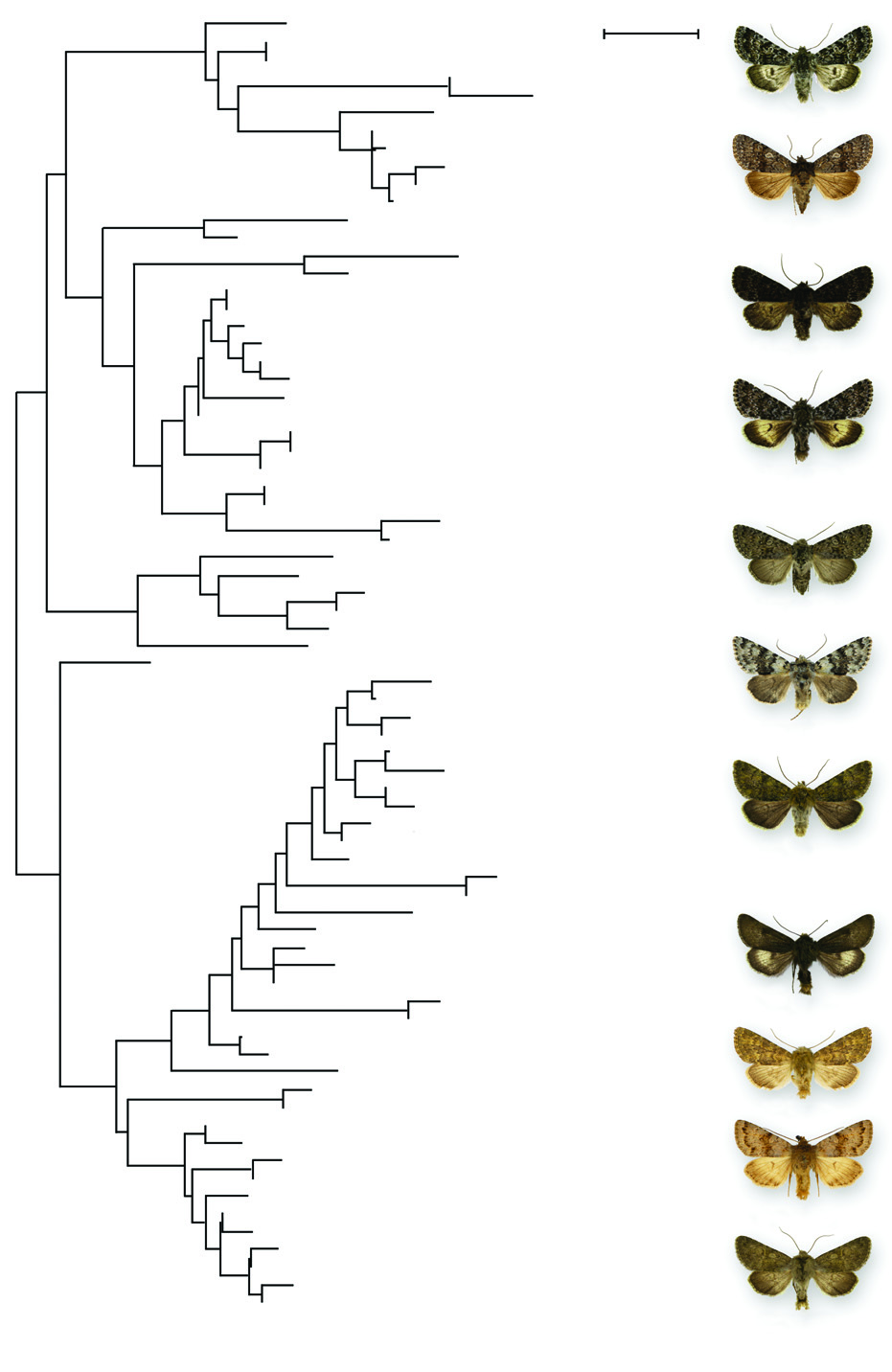
The CO1 DNA barcodes were helpful for this taxonomic revision in several regards. Above the species level DNA was helpful in defining the generic limits. The DNA data support inclusion of L. secedens and exclusion of Lasionhada proxima and the beach species herein transferred to Psammopolia . Within Lasionycta , distance analysis shows species clustering with relatively large differences between such groups supporting the use of species-groups. Th ese longer DNA tree branches mirror the morphology-based species-group classification described in this paper ( Fig. 247 View Figure 247 ). DNA results are less convincing at the sub-group level in the large L. leucocycla species group ( Fig. 248 View Figure 248 ). Th e seven sub-groups are comprised of one to ten superficially and structurally similar species. Most of the sub-group members show a tendency to form DNA clusters, but only the L. subfuscula group consisting of a single species lacks overlap with the other sub-groups. Members of the L. staudingeri sub-group are particularly widely dispersed amongst the entire speciesgroup and its species or species pairs differ by over 6 bp from adjacent species. Although this result could be interpreted to mean that the sub-groups are paraphyletic or polyphyletic, it is probably due to presence of shared haplotypes and non-discrete species haplotype clusters.
Utility of CO1 DNA was also limited at the species level due to the presence of shared haplotypes, close haplotype similarities, and polytypic species as described above. Despite these shortcomings CO1 DNA proved useful in several instances. For example, DNA confirmed that L. brunnea sp. n. from the Canadian Rocky Mountains is distinct from similar-appearing populations of L. uniformis fusca ssp. n. from the central Rocky Mountains. DNA differences support that L. poca , a widely distributed species, and L. frigida sp. n., a closely similar species limited to a small range in the Canadian Rocky Mountains, are distinct, although sympatry of the two species in Alberta was also critical. DNA was also seminal in the decision to treat the disjunct L. poca and L. coloradensis as species rather than subspecies. In the L. perplexa sub-group, large DNA sequence differences first brought to our attention that similar populations from the Pacific Northwest and the central Rocky Mountains, L. perplexella sp. n. and L. subalpina sp. n., were two species.
The distributions of many Lasionycta species reflect survival in Pleistocene glacial refugia and subsequent postglacial dispersal from refugia. Examples of taxa limited to known refugia include L. coracina sp. n., L. carolynae sp. n., and L. subfumosa (Gibson) in Beringia , L. haida sp. n. on the Queen Charlotte Islands of British Columbia, L. uniformis handfieldi in the Gaspé Peninsula of Quebec, and L. leucocycla hampa (Smith) in the White Mountains of New Hampshire. A detailed analysis is beyond the scope of this paper; however, it is interesting to speculate that several distinctive CO1 DNA haplotypes in Pacific Northwest populations of L. subfuscula livida ssp. n. and L. uniformis multicolor ssp. n. could have resulted from montane refugial isolation and postglacial dispersal. If a widespread species became divided into isolated populations during periods of glaciation the isolates would develop different haplotypes due to genetic drift. As a current example, isolated L. subalpina populations in the mountains of northwest Wyoming have distinct DNA haplotypes differing by nearly one per cent from those in the isolated Snowy Range in the southeast part of the state. Blending of populations could occur if the segregates remained interfertile and came into contact after the ice receded. The distinct CO1 haplotypes would be preserved since mitochondrial DNA reflects maternal lineage, whereas other evolved characteristics dependent on nuclear genes would be lost through interbreeding.
Species accounts
The species of Lasionycta are arranged into seven species-groups defined by male and female genital morphology. Th e uncus shape, vesica length, digitus morphology, and cucullus shape are the most important characters for separating species-groups in males. In females, ovipositor lobe shape and vestiture, appendix bursae position and shape, and ductus bursae shape are important distinguishing features. The classification based on morphology is supported by species groupings based on distance analysis of CO1 DNA sequences ( Fig. 247 View Figure 247 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Lasionycta Aurivillius
| Crabo, Lars & Lafontaine, Donald 2009 |
Anartomima
| Boursin C 1952: 55 |
Pseudanarta
| Kozhanchikov I 1947: 18 |
Lasiestra
| Hampson GF 1905: 47 |
Lasionycta
| Aurivillius C 1892: 285 |