
Phyllocnistis maxberryi Kawahara, Nishida & Davis, 2009
|
publication ID |
https://doi.org/ 10.3897/zookeys.27.250 |
|
publication LSID |
lsid:zoobank.org:pub:C6AA8595-6A57-4ACD-B0A1-3AE36F7C8701 |
|
persistent identifier |
https://treatment.plazi.org/id/598E20A2-D76F-41A2-83A0-CA268EC0DF41 |
|
taxon LSID |
lsid:zoobank.org:act:598E20A2-D76F-41A2-83A0-CA268EC0DF41 |
|
treatment provided by |
Plazi |
|
scientific name |
Phyllocnistis maxberryi Kawahara, Nishida & Davis |
| status |
sp. n. |
Phyllocnistis maxberryi Kawahara, Nishida & Davis , sp. n.
urn:lsid:zoobank.org:act:598E20A2-D76F-41A2-83A0-CA268EC0DF41
Diagnosis ( Table 1). Phyllocnistis maxberryi differs from P. drimiphaga and P. tropaeolicola in having an oviform costal fascia with a broad margin, a C-shaped transverse fascia, two costal strigulae, and paired signa that are similar in shape. Unlike drimiphaga and tropaeolicola , the pupa of maxberryi has less developed frontal processes and two parallel rows of spines on the dorsal surface of abdominal segments. Of the three new Phyllocnistis species proposed in this paper, P. maxberryi is morphologically most similar to P. meliacella Becker. Phyllocnistis maxberryi may be distinguished from the latter by its broader apex of the valva and proportionately larger signa.
Adult ( Fig. 2B View Figure 2 ). Forewing length 2.2–3.7 mm. Head. Vestiture silvery white, completely covered with smooth, broad, scales that overlap anterior margin of eye; occipital scales cream. Antenna ̴ equal or slightly longer than length of forewing, scape and pedicel enlarged laterally and covered in long silvery scales, a single row of slender mostly silvery-white scales completely encircling each flagellomere; dorsal surface of antenna with a pale-golden luster. Labial palpus slender, ̴ 0.5 mm in length, with silvery-white scales. Thorax. Forewing silvery white, with a single, broad, light-brown longitudinal fascia with a dark brown posterior margin extending slightly diagonal from base of costa joining costal fascia at ̴ midway to apex; costal fascia oblique, pale gold, oviform, with a broad, inner dark-brown margin; transverse fascia C-shaped, pale gold with dark margin; apical to subapical area pale yellow; two faint, dark-brown costal strigulae present; a single, small black spot at wing apex from which two dark-brown apical strigulae arise. Hindwing silvery white. Legs mostly silvery white, with a faint suffusion of pale gold dorsally over most segments. Abdomen. Length ̴1.5–2.0 mm, silvery white; coremata similar to P. drimiphaga . Male genitalia ( Figs 5 View Figure 5 A–C). Similar to P. drimiphaga except vinculum relatively broader and more U-shaped. Valva ̴ 2× length of vinculum, nearly straight with apex only slightly enlarged ( Fig. 5A View Figure 5 ). Genitalia slide USNM 33279. Female genitalia ( Figs 5 View Figure 5 D–F). Oviscapt greatly reduced as in P. drimiphaga ; ductus bursae completely membranous, slender, elongate, over 12× length of papillae anales and terminating near middle of corpus bursae; corpus bursae greatly enlarged, ̴ 0.7× length of ductus bursae; signa paired, closely similar in shape and size (fusiform), with more posterior signum ̴ 1.2–1.5× longer than anterior signum; each signum with a single, acute, flattened spine projecting from middle ( Fig. 5F View Figure 5 ); length of spines slightly more than width of signa; ductus seminalis extremely slender, elongate, ̴ 1.9× length of corpus bursae and arising from anterior end of corpus bursae. Genitalia slides USNM 33280, 33286.
Larva ( Figs 11 C–F). Mature sap-feeding larva ̴ 6.0 mm long, translucent orange, head capsule brown, prothoracic shield brown ( Figs 10 C–E). Last instar (cocoon-spinning) larva orange, head capsule orange, ̴ 6.3 mm long ( Fig. 10F).
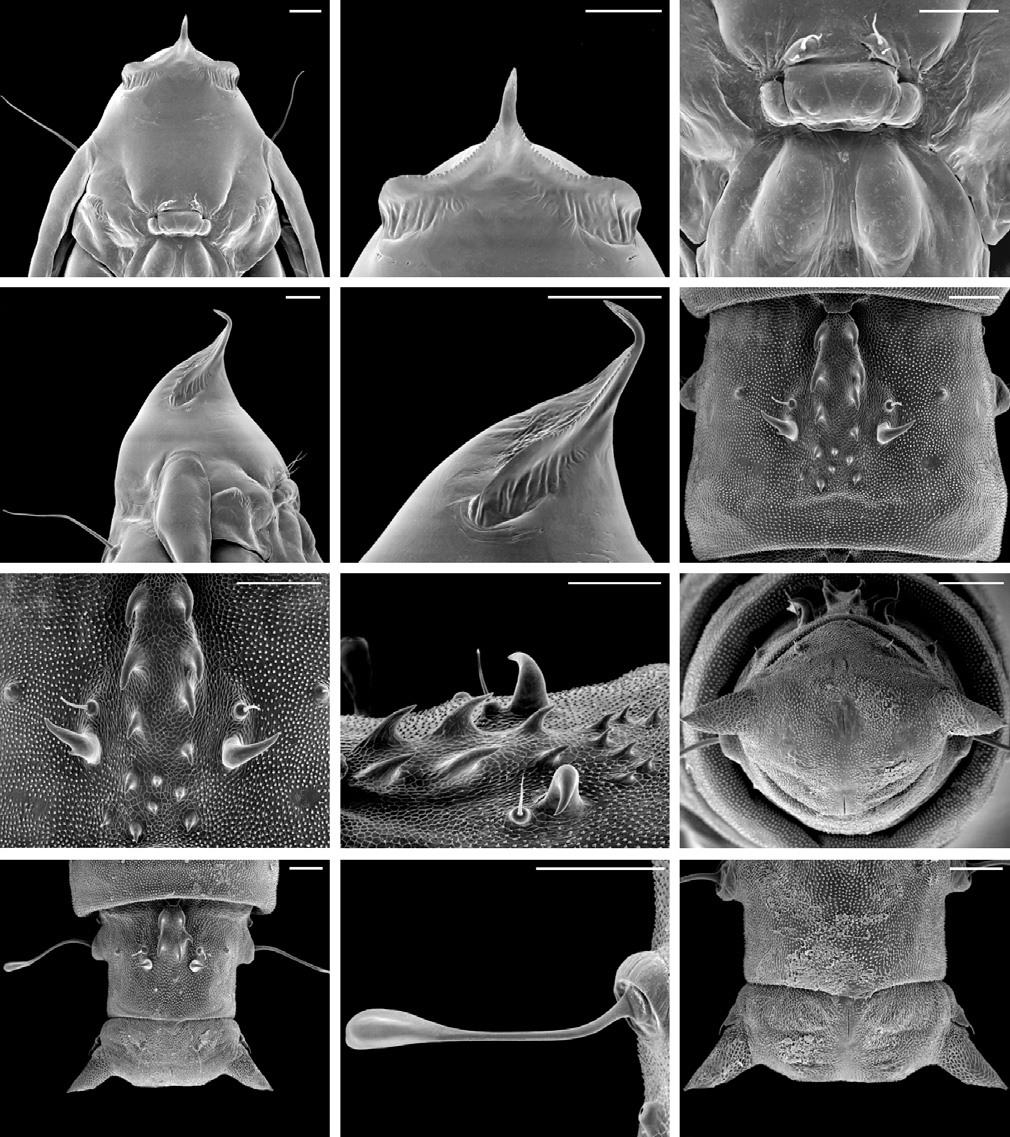
Pupa ( Figs 8 View Figure 8 ; 11H, I). Brown, length up to ̴ 4.0 mm; diameter ̴ 0.85 mm. Vertex with a long, dorsally curved, spine-like process (cocoon-cutter) ( Figs 8A, B, D, E View Figure 8 ), and two pairs of short setae ( Fig. 8C View Figure 8 ). Dorsum of A 2– A 7 with a pair of laterally curved, large spines in between which is a concentration of smaller spines, projecting posteriorly that are roughly arranged in two parallel rows ( Figs 8 View Figure 8 F–H); each segment with a pair of long, lateral, sensory setae ( Fig. 8K View Figure 8 ). A 10 with a pair of slightly divergent processes from caudal apex ( Figs 8I, J, L View Figure 8 ).
Types. Holotype ( Fig. 2B View Figure 2 ): ♀, Costa Rica: Prov. San José, Cerro de la Muerte, Villa Mills, 3100 m, 13 Mar 2003 (adult emergence), host Gaiadendron punctatum , upper leaf miner, col./rear Kenji Nishida, DRD 4474 ( USNM) . Paratypes: Immatures : same locality as holotype: 3 pupae ( USNM 33732 About USNM ), 5 Mar 2003, K. Nishida ; 3 larvae, 2 pupae, 2 Apr 2003, K. Nishida; 1 larva, 21 May 2002, K. Nishida; 3 larvae, 1 pupa ( USNM 34024 About USNM ), 10 Mar 2004, K. Nishida. One pupa, Villa Mills, trail front of La Georgina, 3103 m, 12 Sep 2008, K. Nishida, host Gaiadendron punctatum . Two larvae, 1 pupa, Prov. Heredia, 6 km ENE Vara Blanca, 10°11'N, 84°07'W, 2050 m, 10 May 2005, K. Nishida GoogleMaps ; 1 pupa, 23 Nov 2002, K. Nishida. Adults: same locality as holotype: 1♂, 22 Mar 2003, K. Nishida ; 2♂, 26 Mar 2003, K. Nishida; 2♂, 2♀, Prov. Heredia, 6 km ENE Vara Blanca, 10°11'N, 84°07'W, 1950–2050 m, 2 Feb 2003, K. Nishida GoogleMaps ; 2♂, 9 Apr 2002, 1900 m, emerged 22– 28 Apr 2002, host Gaiadendron punctatum , D. and M. Davis. ♂ slide USNM 33279 About USNM ; ♀ slides USNM 33280 About USNM , 33286 About USNM . One paratype, unknown sex (missing abdomen) at UCR, remaining paratypes at USNM .
Life history ( Fig. 11). Active mines were found on fully open young leaves near the tip of a branch. The smallest leaf with an active mining larva measured 12 × 30 mm. Mines were generally found on young plants about 30 cm to 1.5 m tall, in open fields or along exposed dirt roads or trails. In an open swampy field at the ALAS transect near Vara Blanca, many active mines were found on new leaves on young plants less than 1.5 m tall ( Fig. 11A) and very few active mines were found on larger plants bearing flowers or fruit.
Thirty-six of 42 leaves had mines on the adaxial side and the rest had mines on the abaxial side or on both. Up to three mines were observed on a single leaf. These mines were relatively short, serpentine mines with a brown median frass line that became dark brown as the mine widened ( Fig. 11C).
We recognize a general mining pattern for P. maxberryi : the egg is laid on the mid-vein, near the center of the leaf ( Fig. 11C). After hatching, the larva enters the leaf and mines proximally towards the leaf petiole along the mid-vein and turns toward the leaf apex near or at the leaf petiole and mines along the leaf margin. Before reaching the midpoint along the axis of the leaf, the larva travels inward between the mid-vein and leaf margin and travels towards the leaf apex. After nearing the apex, the larva crosses the mid-vein and begins mining the other half of the leaf in a relatively straight line turning back towards the petiole. Once near the petiole, the larva constructs an oval-shaped chamber and molts within. After molting, the cocoon-spinning instar folds the margin while spinning its cocoon. This pupal fold was typically ̴ 7.0 mm long ( Figs 11B, G). Under rearing conditions, the pupal stage lasts between 21–28 days (n = 7). Five female specimens of Chrysocharis sp. ( Eulophidae : Entedoninae ) were reared from pupal cocoon folds collected at Villa Mills, Cerro de la Muerte.
Host. Gaiadendron punctatum (Ruiz & Pav.) G. Don ( Loranthaceae ) ( Fig. 1G). The free-standing root parasite/epiphyte tree genus Gaiadendron includes approximately 15 species occurring in the New World ( Gentry 1996; Missouri Botanical Garden 2009). Gaiadendron punctatum is distributed from Nicaragua through southern Central America to Bolivia (̴ 17°50'S) between 600 and 4100 m elevation ( INBio 2009; Missouri Botanical Garden 2009). Trees are typically 2–5 m in height with bright yellow/orange flowers ( Kappelle 2008). Young leaves are pale green to reddish brown, about 3–6 cm long and 1–3 cm wide ( KN, pers. obs.). Among species in the genus, only G. punctatum is known from Costa Rica, and it has been recorded above 1500 m in open areas and along trails in cloud forests ( INBio 2009; Kappelle 2008).
Distribution. This species appears to have a greater elevational range than the other two, being found between 1950 and 3100 m. Specimens have been collected from Heredia Province, 6 km ENE of Vara Blanca, in the Cordillera Volcánica Central; and Cartago Province, Cerro de la Muerte, Villa Mills, in Cordillera de Talamanca.
Etymology. Named for the Honorable Max N. Berry of Washington, D.C., an honorary member of the Smithsonian National Board.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |