
Stenoscelida aurantiacus, Müller & Garcia & Fonseca, 2022
|
publication ID |
https://doi.org/ 10.1080/14772019.2022.2128913 |
|
DOI |
https://doi.org/10.5281/zenodo.7331380 |
|
persistent identifier |
https://treatment.plazi.org/id/03C1C248-FFD7-B77E-6FFC-FB5AFBB5B911 |
|
treatment provided by |
Valdenar |
|
scientific name |
Stenoscelida aurantiacus |
| status |
gen. et sp. nov. |
Stenoscelida aurantiacus gen. et sp. nov.
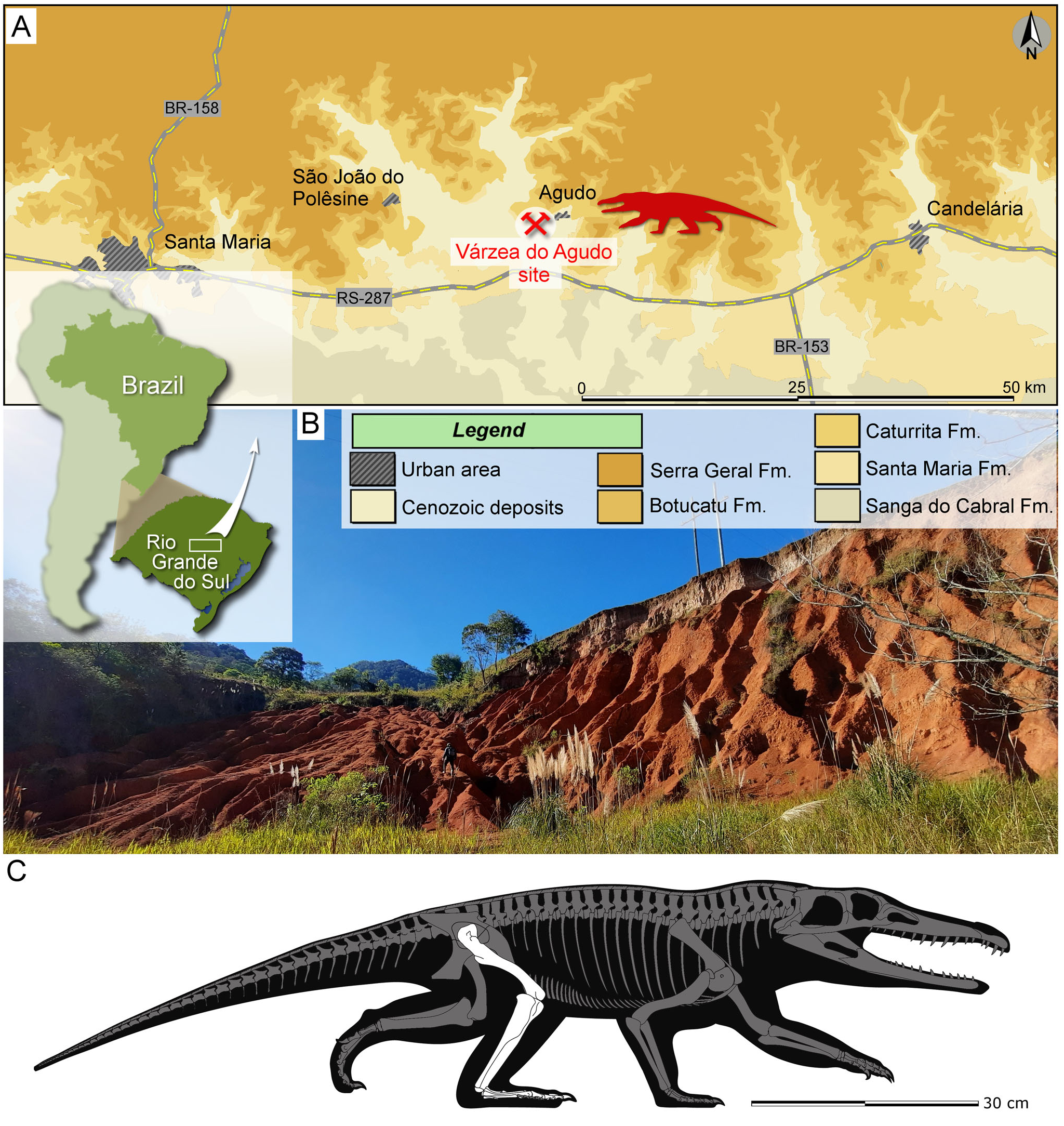
( Figs 1–5 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 )
Holotype. CAPPA/UFSM 0293 , a complete and articulated right hind limb.
Etymology. The genus combines the Greek words UIƐvόç (¼ narrow) and UĸƐƛ o ç (¼ hind leg), referring to the slender leg of the creature. The specific epithet derives from the Latin word aurantiacus (¼ orange), in allusion to the orange colour of the outcropping sediments of the Varzea do Agudo site ( Fig. 1B View Figure 1 ).
Type locality. Varzea do Agudo site (¼ Janner site; 29 ° 39 0 10.89 00 S, 53 ° 17 0 34.20 00 W), Agudo, Rio Grande do Sul, Brazil ( Fig. 1 View Figure 1 ).
Stratigraphic horizon. Lower portion of the Candelaria Sequence ( Horn et al. 2014) of the Santa Maria Supersequence ( Zerfass et al. 2003), Parana Basin. The predominance of the cynodont genus Exaeretodon places the site in the upper part ( Exaeretodon sub-Assemblage Zone; Muller & Garcia 2020) of the Hyperodapedon Assemblage Zone ( Schultz et al. 2020), which is biostratigraphically correlated with the Exaeretodon biozone from the Ischigualasto Formation ( Martınez et al. 2013). A Bayesian age-model for the profile of the Ischigualasto Formation at the Hoyada del Cerro Las Lajas locality in Argentina recovered an age of 227.94 + 0.83 /¯1.67 for the top of the Hyperodapedon AZ ( Desojo et al. 2020), indicating an age of c. 228 Ma for the Exaeretodon biozone. A similar age (late Carnian/early Norian, Late Triassic) for the Exaeretodon sub-Assemblage Zone is inferred in the Candelaria Sequence.
Ontogenetic assessment. The presence of some muscle attachment structures (e.g. anterior trochanter; anterolateral scar) in the femur of CAPPA/UFSM 0293 suggests some degree of development if the ontogenetic pathways of other archosauriforms are considered ( Griffin & Nesbitt 2016). However, it is currently not possible to confirm whether the specimen reached its maximum size or not.
Diagnosis. Stenoscelida aurantiacus differs from all other known proterochampsids in (* local autapomorphies): possessing a slender femur; presence of an anterior trochanter on the femoral head; presence of a raised anterolateral scar above the anterior trochanter; fourth trochanter restricted to the proximal half of the femur; tibia approximately 84% the total length of the femur; in proximal view, the cnemial crest of the tibia is subequal in size to the proximal condyles; fibula with an iliofibularis tubercle on the proximal portion of the shaft *; proximal third of the fibular shaft is anteroposteriorly expanded, tapering distally *; ratio between the minimum midshaft width of metatarsal II and its total length is 0.12; ratio between femoral length and metatarsal III is 2.5; and digit V with a phalanx *.
Description and comparison
Overview
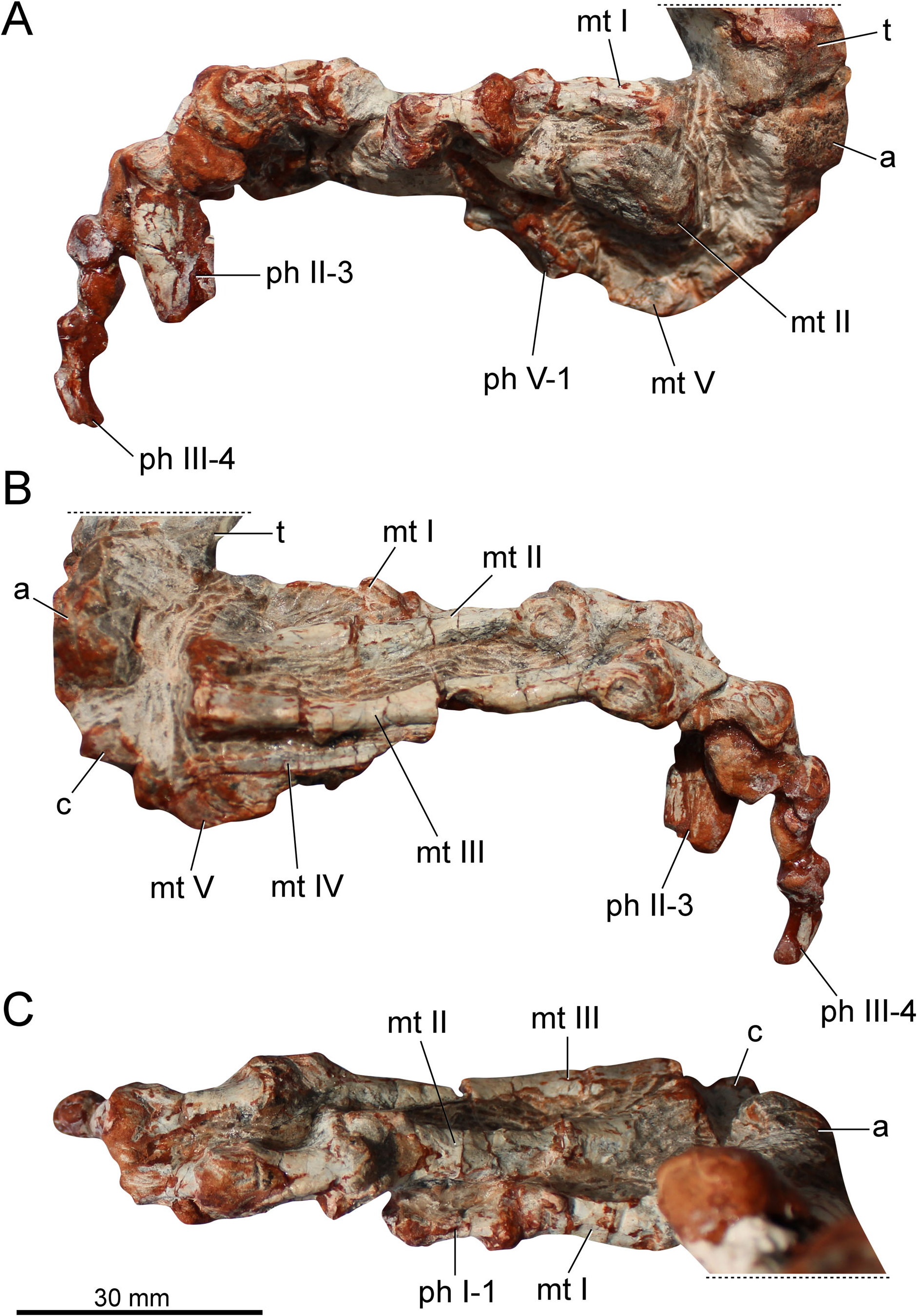
The preserved right hind limb of the holotype is entirely articulated ( Fig. 2 View Figure 2 ). Some portions of the specimen are covered by a thick concretion layer, especially the femoral midshaft and the ankle. The external surface is wellpreserved in several places, revealing some muscle attachment points. However, the specimen was deformed by sedimentary compression. Therefore, the elements are lateromedially compressed, affecting the original shape. As result, the midshaft of the limb bones is collapsed, showing artificial longitudinal grooves or sulci. Part of the fibular midshaft is not preserved. The same is true for the distal portion of metatarsal IV, precluding us from determining its total length. According to the femoral length (147 mm), the specimen is smaller than the Argentinian taxa from the Ischigualasto Formation: Pseudochampsa ischigualastensis (155 mm; Trotteyn & Ezcurra 2014) and Proterochampsa barrionuevoi (179 mm; Trotteyn 2011).
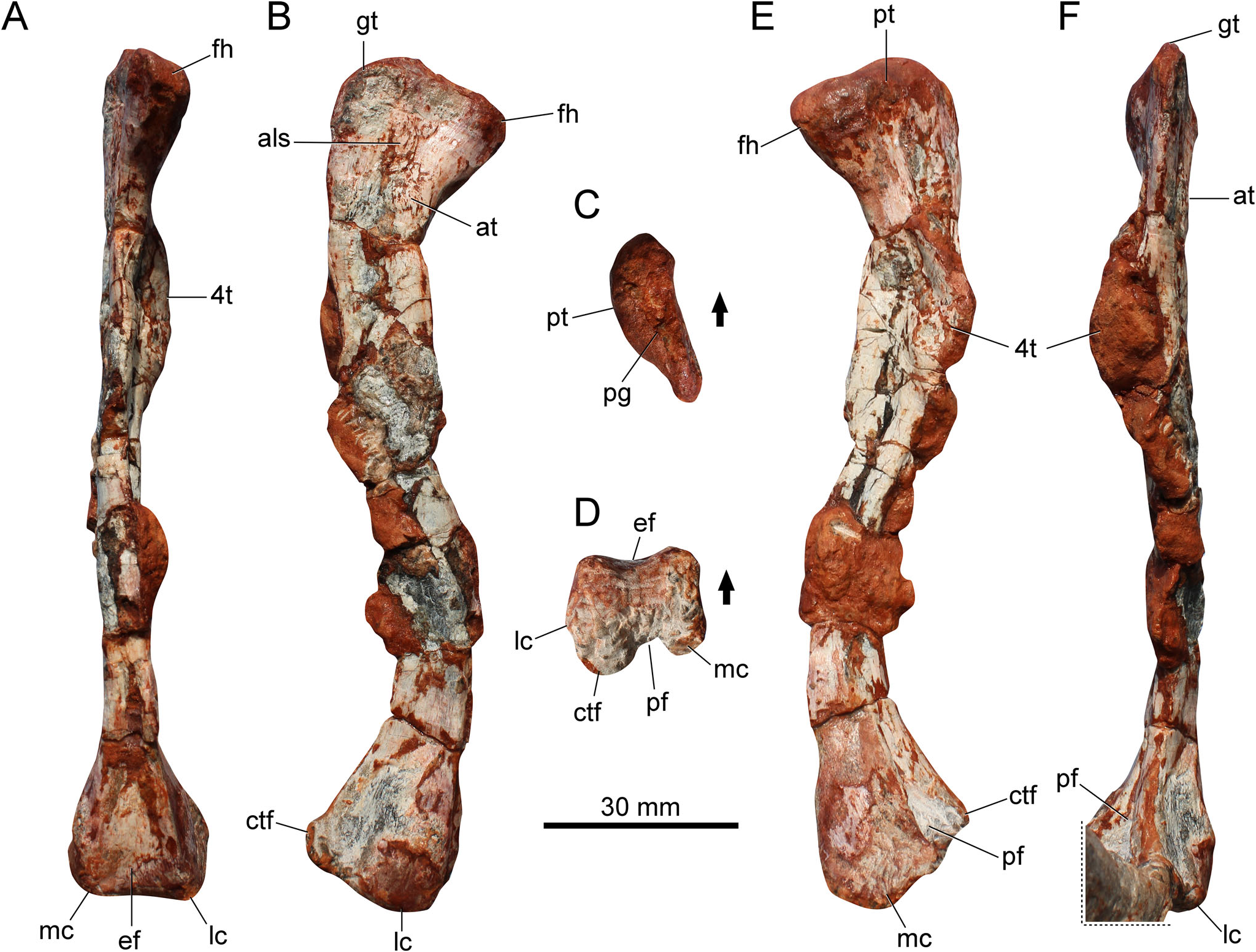
Femur
The femur ( Fig. 3 View Figure 3 ) is sigmoid in lateral and medial views. The extremities are well ossified and expanded. The femoral head is mainly anteriorly directed, whereas in other proterochampsids it is more medially oriented. Diagenetic processes might have exaggerated this condition. The general morphology of the femoral head resembles that of proterochampsids, whereas it differs from the anteromedially expanded femoral head of dinosaurs ( Nesbitt 2011) and from the hook-shaped head of lagerpetids and pterosaurs ( Ezcurra et al. 2020a). The proximal articular surface bears a shallow straight groove ( Fig. 3C View Figure 3 ), which is usually absent in proterochampsids, except in an unnamed rhadinosuchine ( Ezcurra et al. 2019; CRILAR-Pv 491) from the Chanares Formation. There is an anterior tuber (sensu Ezcurra 2016) on the proximal portion of the femur, and although the bone lacks a posteromedial tuber, the posterior tuber is present. The greater trochanter is rounded and tall ( Fig. 3B View Figure 3 ), differing from the typical angled trochanter of dinosaurs, and in the same way, there is no dorsolateral trochanter on the proximal portion of the bone. Conversely, the anterolateral surface bears a raised and rugose area ( Fig. 3B View Figure 3 ) where the anterior (¼ lesser) trochanter is reported for a number of archosauriforms (e.g. ornithosuchids, aetosaurs, dinosauriforms). Whereas the anterior trochanter is absent in several proterochampsids ( Trotteyn et al. 2013), the structure occurs in the holotype of Gualosuchus reigi (PULR-V 05; M. D. Ezcurra, pers. comm.). A trochanteric shelf is not associated with the anterior trochanter, and the proximal-most portion of this trochanter is completely connected to the shaft. Slightly above the proximal tip of the anterior trochanter there is an additional scar, which resembles the anterolateral scar (sensu Griffin & Nesbitt 2016) of aphanosaurs and dinosauriforms. There is a similar scar in Gualosuchus reigi (PULR-V 05; M. D. Ezcurra, pers. comm.).
The transition from the femoral head to the shaft is smooth. The fourth trochanter rests on the posterior surface of the proximal third of the femur ( Fig. 3A View Figure 3 ). It is crest-like and symmetrical in shape. Its medial surface is ornamented with muscle scars. The distal portion of the fourth trochanter does not extend further down along the femoral shaft. In contrast, the fourth trochanter is strongly proximodistally developed in Gualosuchus reigi and Chanaresuchus bonapartei ( Ezcurra et al. 2019) . The midshaft is slender. The robustness index (RI, sensu Wilson & Upchurch 2003; i.e. average of the greatest widths of the proximal end, midshaft and distal end of the element divided by the length of the element) is 0.14. For comparison, this is slightly slenderer than that of some early dinosaurs, such as Gnathovorax cabreirai (0.16) and Buriolestes schultzi (0.15; Muller & Garcia 2022).
The anterior surface of the distal portion bears an extensor fossa ( Fig. 3D View Figure 3 ), which results in a concave anterior margin in distal view. On the opposite side, the popliteal fossa is well delimited ( Fig. 3E View Figure 3 ). It is proximodistally short, approximately 10% of the total length of the bone. This condition distinguishes the specimen from silesaurids and aphanosaurs, where the fossa is considerably longer ( Nesbitt et al. 2010, 2017). The distal condyles are approximately at the same level in anterior or posterior views. In addition, the lateral surface between the lateral condyle and the crista tibiofibularis is smooth, lacking a deep groove.
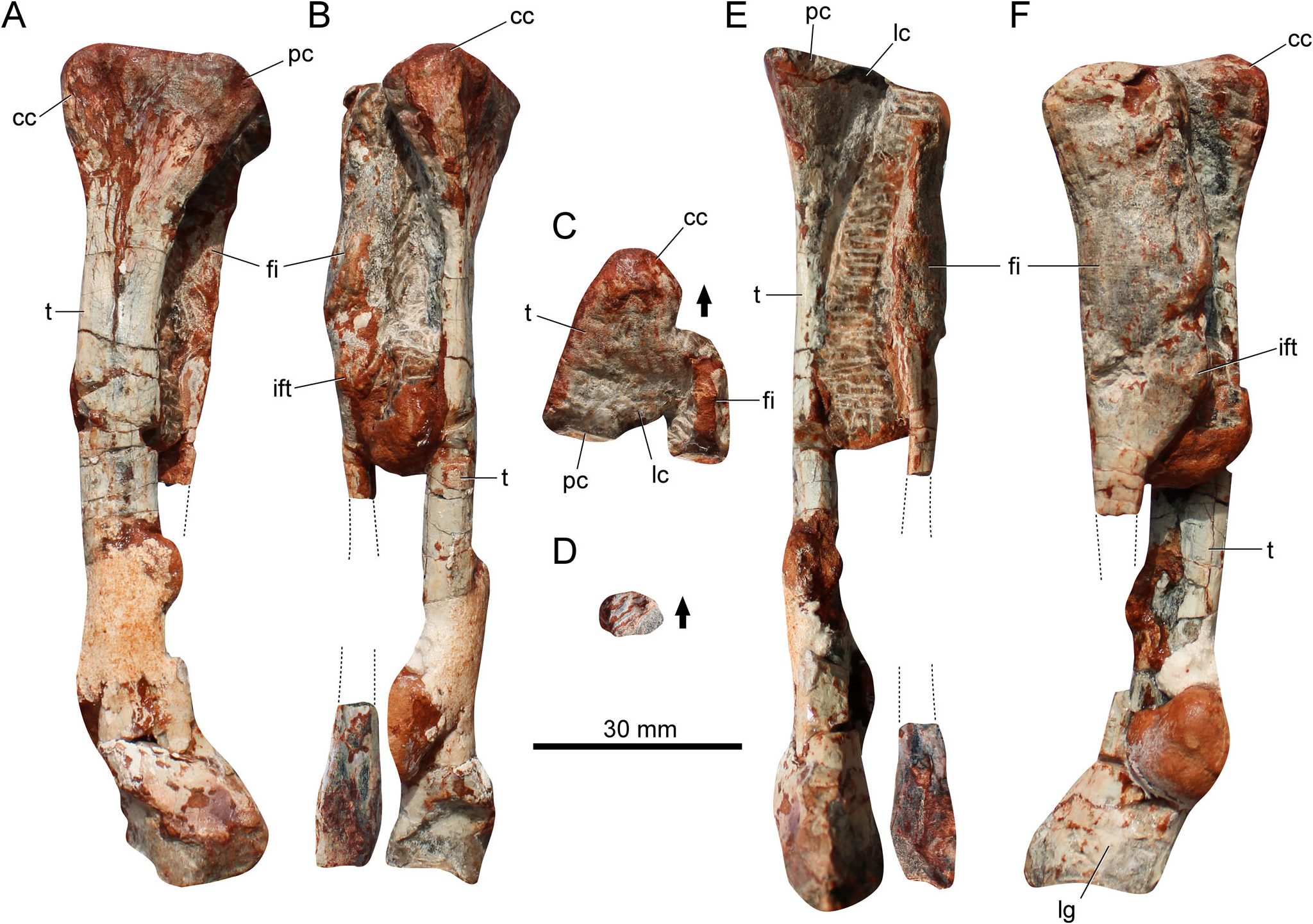
Tibia
The tibia ( Fig. 4 View Figure 4 ) is 84% the total length of the femur ( Table 1 View Table 1 ). This resembles the condition in Pseudochampsa ischigualastensis , where it is approximately 82% ( Trotteyn & Ezcurra 2014), and differs from both Proterochampsa barrionuevoi (74.5%; Trotteyn 2011) and Tropidosuchus romeri (100%; Arcucci 1990). The bone is straight in anterior/posterior or medial/lateral views. The proximal portion is strongly expanded and is triangular in shape in proximal view ( Fig. 4C View Figure 4 ). The anterior half of the proximal margin is proximally projected in medial view. In medial or lateral views, the well-developed cnemial crest extends anteriorly, and in proximal view it is straight and the anterior margin is rounded. The cnemial crest is sub-equal in size regarding the proximal condyles, distinguishing the new species from Proterochampsa barrionuevoi , where the crest is proportionally smaller. The posterior and lateral condyles are sub-equal in size. The lateral condyle is offset regarding the posterior condyle. The posterior condyle does not taper posteriorly. The presence of any crest on the lateral surface of the proximal portion of the bone is uncertain.
The midshaft is ovoid in cross-section. The bone wall is thick, distinct from the thin walls of several pan-avians and some crocodylomorphs ( Kellner et al. 2022). The distal portion of the bone is moderately expanded. It lacks a posterolateral process. There is a proximodistally oriented groove on the lateral surface of the distal portion ( Fig. 4F View Figure 4 ). It is more pronounced in Proterochampsa barrionuevoi ( Trotteyn 2011) . The distal outline of the tibia is elliptical. It is anteroposteriorly longer than transversely wide.
·
Fibula
The total length of the fibula ( Fig. 4 View Figure 4 ) is uncertain because a portion of the midshaft is not preserved. The bone is straight in anterior or lateral views, and the proximal articular surface is anteroposteriorly expanded and transversely compressed ( Fig. 4C View Figure 4 ). It differs from the rounded to elliptical proximal end of some pseudosuchians (e.g. Dynamosuchus collisensis , Prestosuchus chiniquensis ). In proximal view, the lateral margin is convex but the medial margin is concave. In lateral view, the posterior margin projects posteriorly, whereas the anterior margin lacks any anterior expansion. Conversely, in non-proterochampsid proterochampsians (e.g. Vancleavea campi ; Litorosuchus somnii ; Jaxtasuchus salomoni ) the proximal portion is symmetrical to nearly symmetrical in lateral view. The posterior margin of the proximal third of the bone is sharp.
The proximal third of the midshaft is anteroposteriorly expanded, being wider than the tibia ( Fig. 4F View Figure 4 ). Conversely, the shaft becomes extremely slender for the next two-thirds, being approximately two times narrower than the tibia. This is an unusual condition and distinguishes the specimen from other proterochampsids. Furthermore, there is an iliofibularis tubercle on the proximal portion of the shaft ( Fig. 4E, F View Figure 4 ), a feature absent in other proterochampsids. The condition in Stenoscelida aurantiacus differs from the more distally located tubercle of several pseudosuchians and rhynchosaurs.
The distal portion of the fibula is gently expanded. It is approximately 0.45 times the maximum length of the proximal articular surface ( Table 1 View Table 1 ). There is a faint longitudinal ridge running on the anterolateral margin. The distal articular surface is flat to concave. In distal view ( Fig. 4D View Figure 4 ), it is ovoid to triangular.
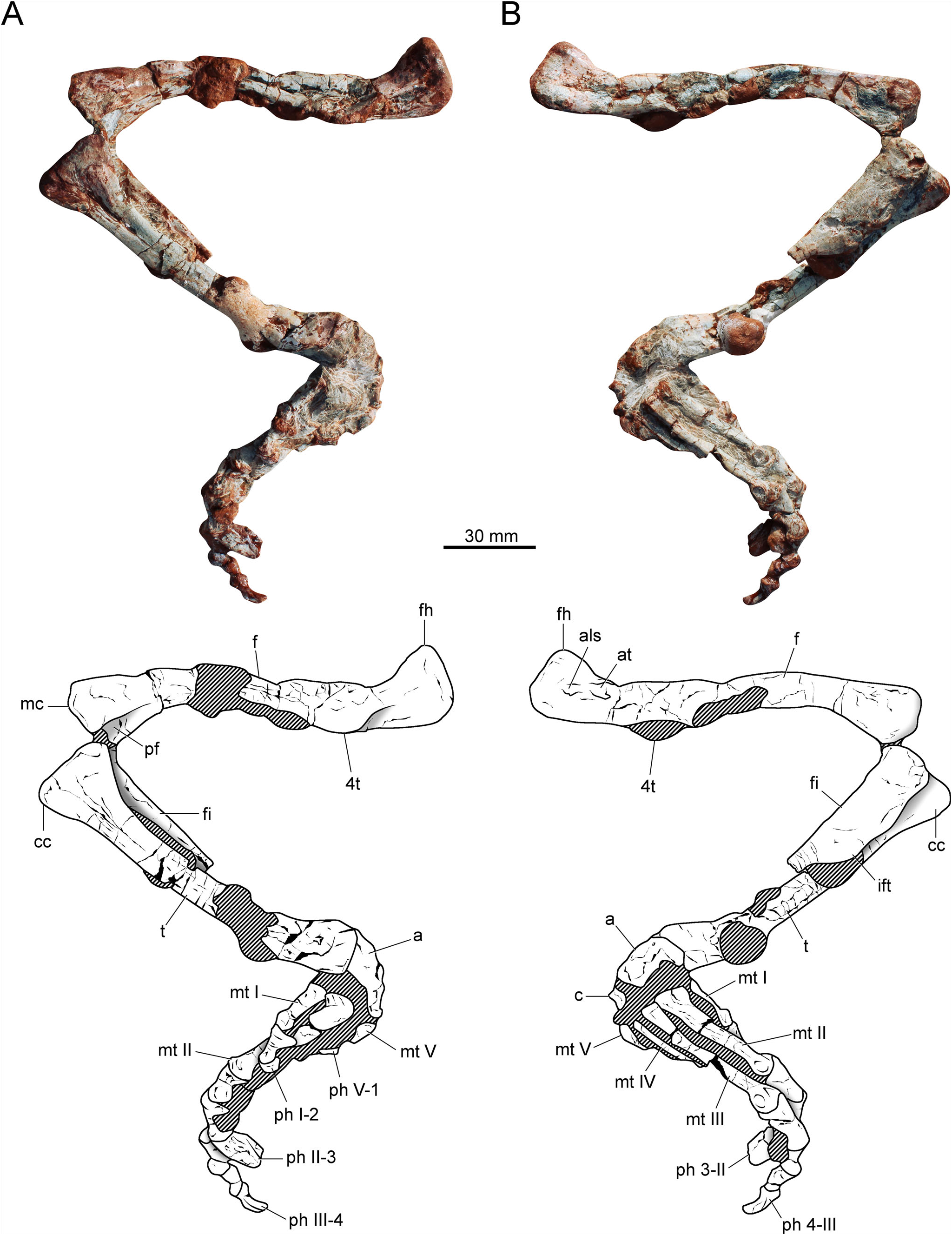
Proximal tarsals
The astragalus and calcaneum are poorly preserved ( Figs 2 View Figure 2 , 5). The astragalus is transversely wider (20.5 mm) than anteroposteriorly long (11 mm). The anterior margin of the astragalus is concave in dorsal view ( Figs 2B View Figure 2 , 5B). A transverse groove runs on the posterior surface of the bone. The presence or absence of foramina on this posterior groove is uncertain because it is badly preserved. As in other proterochampsids, the tibial facet is wider than the fibular facet. The latter facet is dorsolaterally oriented, resembling the condition of Chanaresuchus bonapartei and Proterochampsa barrionuevoi , whereas in Pseudochampsa ischigualastensis it is dorsally oriented ( Trotteyn & Ezcurra 2014). The ventral surface of the bone is transversely convex. The calcaneum is 10 mm in width (approximately half of the astragalar width). There is a posterolaterally oriented calcaneal tuber. This structure is sub-rectangular, with a straight lateral margin. In dorsal view, the posterolateral corner of the calcaneal tuber is rounded ( Fig. 5B View Figure 5 ).
Metatarsals
As in other proterochampsids, the metatarsals ( Fig. 5 View Figure 5 ) overlap each other. Metatarsal I is shorter than metatarsals II–IV ( Fig. 5C View Figure 5 ). It is almost three times shorter than metatarsal III ( Table 1 View Table 1 ). The proximal articular surface is transversely expanded. There is a longitudinal smooth crest on the dorsomedial corner of the proximal half of the bone. The distal end is moderately expanded. The lateral condyle is more pronounced dorsally than the medial one. There is a shallow extensor fossa on the dorsal surface of the distal portion. The presence of collateral ligament pits on the condylar sides is uncertain. The distal articular surface is not strongly ginglymoid.
Metatarsal II is the stoutest metatarsal. The proximal articular surface is strongly expanded ( Table 1 View Table 1 ). The ratio between the proximal articular surface and the total length is 0.39. It is 0.36 in Rhadinosuchus gracilis ( Ezcurra et al. 2015) . Conversely, the shaft is stouter in Rhadinosuchus gracilis , where the ratio between the minimum midshaft width and the total length of metatarsal II is 0.17. In Stenoscelida aurantiacus it is 0.12. On the dorsal surface of the distal portion, there is a triangular extensor fossa. It is not possible to determine if the articular surface is ginglymoid. There is no evidence of a collateral ligament pit on the medial surface of the medial condyle. On the other hand, the lateral surface of the lateral condyle bears a well-delimited collateral ligament pit ( Fig. 5B View Figure 5 ). In dorsal view, the lateral condyle is straight, whereas the medial condyle is slightly medially oriented. Both condyles are equally expanded distally, which is distinct from Rhadinosuchus gracilis , where the lateral condyle is far more extended distally ( Ezcurra et al. 2015).
Metatarsal III is the longest ( Table 1 View Table 1 ), whereas its midshaft is slenderer than that of metatarsal II. Its proximal articular surface is moderately expanded (maximum proximal width is 14 mm), far less than the proximal articular surface of metatarsal II (19 mm). The distal articular surface resembles that of metatarsal II, with a marked extensor fossa on the dorsal surface and a collateral ligament pit on the lateral surface of the lateral condyle ( Fig. 5B View Figure 5 ). The presence of the collateral ligament pit on the medial condyle is uncertain (it is covered by thick sediment). The lateral condyle is slightly more pronounced laterally than the lateral condyle of metatarsal II.
Metatarsal IV is incomplete ( Fig. 5B View Figure 5 ). The element does not preserve the distal portion. Therefore, its total length is uncertain. The preserved part is 30 mm in length. Despite its condition, the element is longer than metatarsals I and V. The proximal articular surface is not transversely expanded. The general morphology of the element is simple. It comprises an elongated and compressed shaft. Distinct from the previous metatarsals, metatarsal IV is plate-like in cross-section.
Metatarsal V is the shortest ( Fig. 5B View Figure 5 ), being slightly shorter than phalanx 1 of digit I. In this sense, Stenoscelida aurantiacus differs from Pseudochampsa ischigualastensis , where metatarsal V is slightly longer than the phalanx 1 from digit I ( Trotteyn & Ezcurra 2014). The proximal articular surface is moderately expanded and ‘U’-shaped in ventral view. The shaft tapers distally to an unexpanded and featureless distal end, which bears no distal condyles.
Phalanges
The phalangeal formula is 2-3-4-?-1 ( Fig. 5 View Figure 5 ). Other proterochampsids lack phalanges in digit V. The first phalanx of digit I is longer than broad. This is the same pattern observed in the other phalanges of the specimen. This phalanx is shorter than the first phalanx from digits II and III ( Table 2 View Table 2 ). The dorsal intercondylar process is moderately developed and the distal articular surface is ginglymoid. There is an extensor depression on the dorsal surface of the distal portion. This surface receives the extensor tubercle from the ungual phalanx. The ungual (distal-most) phalanx of digit I lacks its distal half. The phalanx is transversely compressed and tapers distally (at least, the preserved portion). The flexor tubercle is poorly developed ( Fig. 5A View Figure 5 ). The first phalanx of digit II is similar in shape to that of digit I. However, the phalanx from digit II is larger ( Fig. 5C View Figure 5 ). Phalanx 2 is slightly smaller. Its anatomy is obscured by a thick layer of concretion. The ungual phalanx of digit II is clearly the largest ungual. It is transversely compressed and tapers to a sharp point. The ungual is not ventrally recurved and lacks a well-developed flexor tubercle. Digit III is the longest ( Fig. 5B View Figure 5 ). The first phalanx from digit III is sub-equal in length to the first phalanx of digit II ( Table 2 View Table 2 ). On the other hand, the shaft of this phalanx is more gracile. The extensor fossa is deep on the dorsal surface of the distal portion. The subsequent phalanges of this digit are smaller. The ungual is remarkably smaller than that of digit II. The number of phalanges in digit IV is unknown and the distal portion of metatarsal IV is not preserved. Therefore, the absence of phalanges is ambiguous. The phalanx of digit V ( Fig. 5A View Figure 5 ) is reduced to an elongated structure with a concave proximal articular surface. No condyles or other structures are present in this vestigial element.
Phylogenetic results
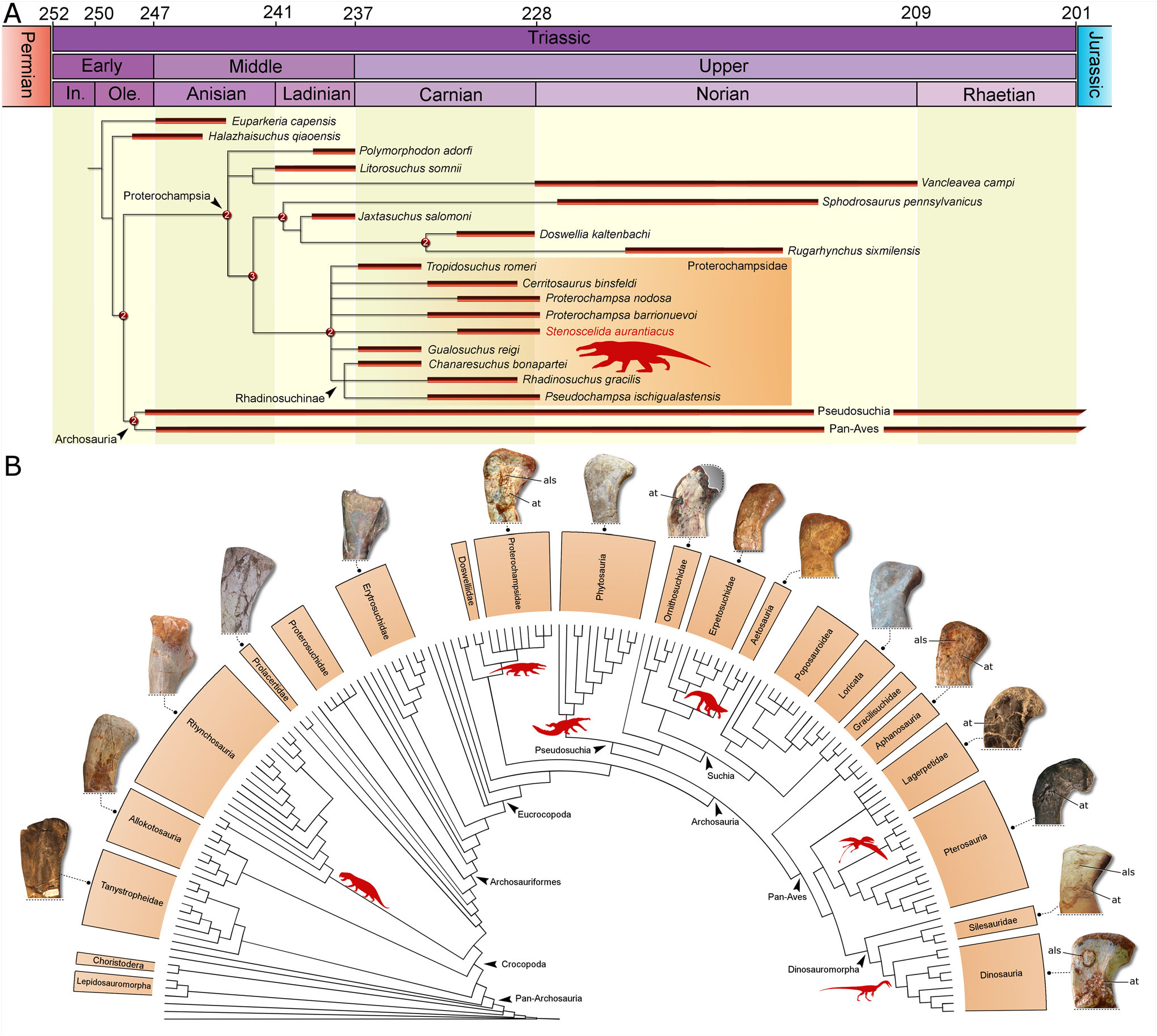
The phylogenetic analysis recovered 39,200 most parsimonious trees (MPTs) of 6287 steps (consistency index ¼ 0.18375; retention index ¼ 0.63457). Stenoscelida aurantiacus nests within Proterochampsidae in all MPTs ( Fig. 6A View Figure 6 ). The clade is supported by 19 synapomorphies, of which three are coded for Stenoscelida aurantiacus : 1) midshaft diameter of metatarsal II is more than the midshaft diameter of metatarsal I (ch. 572: 0! 1); 2) midshaft diameter of metatarsal IV is lower than that of metatarsal III (ch. 573: 0! 1); and 3) crest-like fourth trochanter, dorsoventrally/anteroposteriorly taller than or equal to the shaft at its minimum depth (ch. 803: 1! 2). Proterochampsidae is the sister group to Doswelliidae , which is composed of Rugarhynchus sixmilensis, Sphodrosaurus pennsylvanicus , Doswellia kaltenbachi and Jaxtasuchus salomoni . The clade supporting Proterochampsidae + Doswelliidae is in a trichotomy with Polymorphodon adorfi and an unnamed clade composed of Vancleavea campi and Litorosuchus somnii . These clades are included within Proterochampsia.
Regarding the internal relationships of Proterochampsidae , there is a polytomy composed of Stenoscelida aurantiacus , Tropidosuchus romeri , Cerritosaurus binsfeldi , Gualosuchus reigi , both species of Proterochampsa , and Rhadinosuchinae in the strict consensus tree ( Fig. 6A View Figure 6 ). Given the absence of overlapping elements between the new taxon and several other proterochampsids (e.g. Proterochampsa nodosa , Cerritosaurus binsfeldi ), these unresolved affinities are unsurprising. The new taxon, however, does not nest within Rhadinosuchinae in any of the MPTs, thus Stenoscelida aurantiacus is a non-rhadinosuchine proterochampsid. The clade Rhadinosuchinae is composed of a trichotomy including Rhadinosuchus gracilis , Pseudochampsa ischigualastensis and Chanaresuchus bonapartei . Except for the polytomy at the base of Proterochampsidae , the topology of the strict consensus tree is identical to that recovered by Ezcurra & Sues (2021).
Discussion
Taxonomic status
In addition to a series of cranial features, proterochampsids are characterized by a metatarsal II with a robust midshaft, whereas metatarsal IV has a slender profile ( Trotteyn et al. 2013). The new taxon possesses this particular combination of traits. Therefore, Stenoscelida aurantiacus is unequivocally assigned to Proterochampsidae due to the shape of metatarsals. Moreover, this is supported by our phylogenetic analysis. Unfortunately, the absence of overlapping elements between Stenoscelida aurantiacus and several other proterochampsids (especially the Brazilian forms) precludes a clear assessment of its specific phylogenetic relationships. Rhadinosuchus gracilis is the only Brazilian proterochampsid from the Hyperodapedon AZ that preserves any hind limb elements. Actually, the holotype preserves a left metatarsal II ( Ezcurra et al. 2015). It is the only comparable element between both taxa. The latter is a member of Rhadinosuchinae ( Ezcurra et al. 2015), a less inclusive clade within Proterochampsidae , a nesting reinforced by our results (see above). On the other hand, Stenoscelida aurantiacus is not recovered as a rhadinosuchine in our analysis, and despite the few comparable elements, there are some differences between the metatarsal II of CAPPA/ UFSM 0293 ( Stenoscelida aurantiacus ) and SNSB- BSPG AS XXV 50 ( Rhadinosuchus gracilis ). In SNSB- BSPG AS XXV 50 the midshaft is stouter and the proximal articular surface is proportionally smaller than in CAPPA/UFSM 0293. The medial and lateral condyles of SNSB-BSPG AS XXV 50 are uneven distally, whereas in CAPPA/UFSM 0293 both condyles are in the same plane. A more detailed comparison demands additional overlapping elements. In sum, CAPPA/UFSM 0293 cannot be ascribed to Rhadinosuchus gracilis according to the lack of anatomical correspondence and distinct phylogenetic positions.
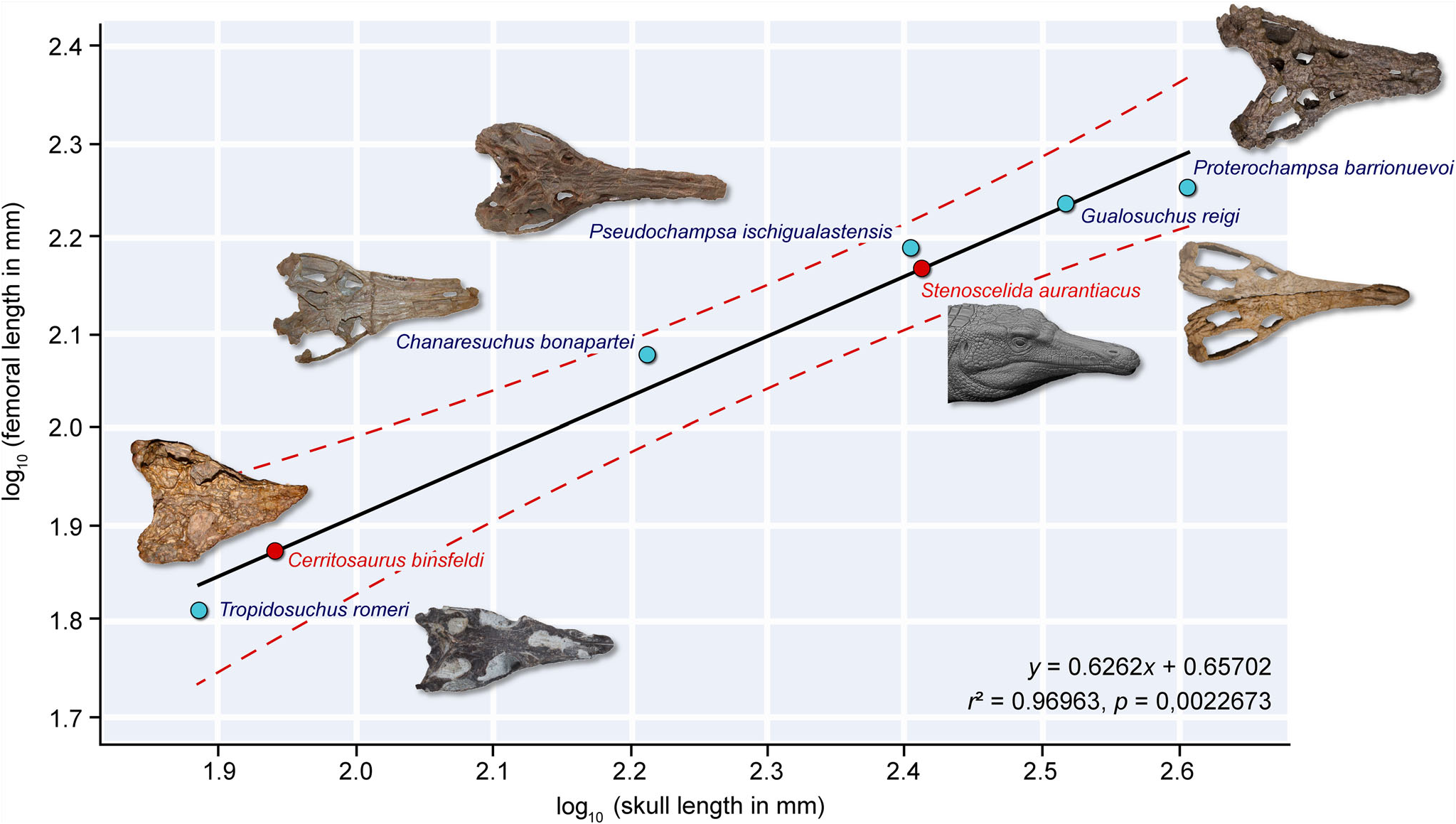
Although the holotype (CA unnumbered) of Cerritosaurus binsfeldi preserves some postcranial elements, there are no hind limb bones ( Price 1946; Trotteyn et al. 2013). Despite the absence of overlapping bones between Cerritosaurus binsfeldi and Stenoscelida aurantiacus , the former is far smaller, with a cranial length of 8.9 cm ( Trotteyn et al. 2013). For comparison, the skull and femoral lengths of other proterochampsids are, respectively: Chanaresuchus bonapartei , 16.3 cm and 11.96 cm ( Trotteyn & Ezcurra 2020); Gualosuchus reigi , 33 cm and 17.2 cm ( Ezcurra et al. 2020b); Proterochampsa barrionuevoi , 40.45 cm and 17.9 cm ( Trotteyn & Haro 2011); Pseudochampsa ischigualastensis , 25.4 cm and 15.48 cm ( Trotteyn & Ezcurra 2014); and Tropidosuchus romeri , 7.67 cm and 6.46 cm ( Bestwick et al. 2022). Based on these measurements, the estimated femoral length of Cerritosaurus binsfeldi is 7.54 cm (y ¼ 0.6262 x + 0.65702; Fig. 7 View Figure 7 ), which is approximately two times shorter than the femoral length of Stenoscelida aurantiacus (i.e. 14.7 cm). So far, there are no ontogenetic assessments regarding Cerritosaurus binsfeldi . Therefore, additional studies or new fossils are necessary in order to determine if the size discrepancy between both species is an effect of ontogeny or another intraspecific factor. Nevertheless, geological and stratigraphical evidence provides further support for our taxonomic proposal. Cerritosaurus binsfeldi was excavated from strata ascribed to the Hyperodapedon Acme Zone ( Langer et al. 2007), which are older than that which yielded Stenoscelida aurantiacus (i.e. Exaeretodon sub-AZ; Langer et al. 2007; Martınez et al. 2013; Desojo et al. 2020; Muller & Garcia 2020; Schultz et al. 2020).
In relation to the occurrence of Proterochampsa nodosa in sediments attributable to the Exaeretodon sub-AZ, Stenoscelida aurantiacus does not present overlapping elements with the former, which is known from a single skull with mandible in occlusion ( Barberena 1982; Trotteyn et al. 2013; Sim ~ ao-Oliveira et al. 2022). Although there are no comparable elements between both Brazilian specimens, it is possible to compare Stenoscelida aurantiacus with the Argentinian species of the same genus: Proterochampsa barrionuevoi . The latter is known from several specimens, including an almost complete skeleton (PVSJ 606), and its hind limb morphology differs substantially from that in Stenoscelida aurantiacus , not to mention the three local apomorphies of the new taxon (see Diagnosis, above), which will not be repeated here. In relation to the femur, the popliteal fossa of CAPPA/UFSM 0293 ( Stenoscelida aurantiacus ) is short, comprising 10% of the total femoral length, but in PVSJ 606 ( Proterochampsa barrionuevoi ; Trotteyn 2011), it is longer in comparison, reaching roughly 23% of the total femoral length. Furthermore, based on femoral measurements, CAPPA/UFSM 0293 is 22% smaller than PVSJ 606, and although it is possible to argue that the specimen probably does not represent an immature individual due to the presence of well-developed femoral epiphyses and scars (see above), we cannot determine with the evidence at hand if CAPPA/UFSM 0293 reached its maximum size. The proportions of the tibia and femur also differ between both specimens, where in CAPPA/UFSM 0293 the tibia is proportionally longer than in PVSJ 606; although we recognize that the size difference between specimens could possibly influence this trait, there is no evidence to support this sort of allometric discrepancy in proterochampsids. Therefore, as in the case of Cerrritosaurus bindfeldi, more specimens are needed in order to test this assumption. Regarding the tibia, in CAPPA/UFSM 0293 the proximal extremity of the tibia is perpendicular in relation to its long axis, whereas in PVSJ 606 it is oblique. Moreover, in PVSJ 606, the cnemial crest is smaller than the proximal condyles ( Trotteyn 2011), whereas in CAPPA/UFSM 0293 it is sub-equal in size to the proximal condyles. The fibula of PVSJ 606 maintains its anteroposterior width for most of its proximodistal length, as in other proterochampsids (although the epiphyses are slightly expanded), but in CAPPA/UFSM 0293 the fibula is significantly expanded in the proximal third of the element and then tapers distally, becoming remarkably narrow, ending in an unexpanded distal epiphysis. In sum, we consider it unlikely that CAPPA/UFSM 0293 represents a new specimen of the genus Proterochampsa .
Distribution of hind limb traits
The occurrence of a new species of proterochampsid with a well-preserved hind limb is particularly exciting. According to several authors (e.g. Sereno 1991; Ezcurra 2016; Ezcurra et al. 2020a; Trotteyn & Ezcurra 2020; Kellner et al. 2022), Proterochampsia is regarded as the sister group to Archosauria. As a result, the anatomy of proterochampsians is crucial in order to optimize the origin and distribution of morphological traits usually present in the earliest diverging clades of archosaurs. Several of these traits rely on the pelvic and hind limb anatomy ( Nesbitt 2011; Ezcurra 2016; Ezcurra et al. 2020a). Ankle anatomy has been historically used to classify Pseudosuchia and Pan-Aves ( Cruickshank 1979), and the presence and shape of trochanters, condyles and other features from the zeugopodium and stylopodium have supported internal relationships among these clades ( Nesbitt 2011; Cabreira et al. 2016; Baron et al. 2017; Ezcurra et al. 2020a). For instance, the anterior (¼ lesser) trochanter is exhaustively reported for the femur of dinosaurs and related groups ( Hutchinson 2001; Pintore et al. 2021). It is treated as the scar of the M. iliofemoralis ( Hutchinson 2001). The shape of the anterior trochanter varies within the main lineages of dinosaurs (Pintore et al. 2021), providing data for phylogenetic analyses ( Nesbitt 2011; Langer & Ferigolo 2013; Baron et al. 2017). Yet, the occurrence of the structure is not restricted to the pan-avian lineage, since the anterior trochanter is reported for crocodylomorphs and ornithosuchids ( Clark & Sues 2002; Baczko et al. 2019; Muller et al. 2020a), which are pseudosuchians. So far, there are no records of the anterior trochanter for non-archosaurian archosaurs (to our knowledge). Stenoscelida aurantiacus (together with Gualosuchus reigi ) provides the first evidence of the presence of this feature in a non-archosaurian archosauriform ( Fig. 6B View Figure 6 ). The occurrence of the anterior trochanter in a proterochampsid supports a deeper origin for this trait than previously thought. It is possible that instead of evolving independently in the two main lineages of Archosauria, the anterior trochanter originated earlier and was retained in the major clades of Pan-Aves and in some members of Pseudosuchia. However, the absence of this trait in the proterochampsian Vancleavea campi cast doubts on this hypothesis and favours a scenario where the anterior trochanter evolved independently in proterochampsids and archosaurs.
Above the anterior trochanter, the femur of Stenoscelida aurantiacus displays another interesting feature. It comprises a raised surface that resembles the anterolateral scar (¼ ‘dorsolateral ossification’ of Piechowski et al. 2014) reported for dinosauriforms and aphanosaurs, both of which are members of Pan-Aves ( Piechowski et al. 2014; Griffin & Nesbitt 2016; Muller 2022). Conversely, the structure is not recognized in pseudosuchians or non-archosaurian archosauriforms ( Fig. 6B View Figure 6 ). It was hypothesized as the ossification of the iliofemoral ligament insertion ( Griffin & Nesbitt 2016). The occurrence of this trait in a non-archosaurian archosauriform implies a similar origin as that of the anterior trochanter, meaning that it could have been secondarily lost in pseudosuchians, whereas in Pan-Aves it was retained. The alternative scenario implies the independent origin of the scar in Pan-Aves and proterochampsids (considering the presence in both Stenoscelida aurantiacus and Gualosuchus reigi ). It has been demonstrated that the presence and shape of the anterolateral scar is affected by intraspecific variation ( Piechowski et al. 2014; Griffin & Nesbitt 2016; Muller 2022). Therefore, the presence/ absence of this trait must be carefully considered.
The occurrence of the iliofibularis tubercle on the fibula of Stenoscelida aurantiacus is another unusual trait for Proterochampsia. Nevertheless, this feature is widely distributed within crocopodans ( Ezcurra et al. 2020a), and it is not regarded as a traditional synapomorphy of any exclusive clade (e.g. Nesbitt 2011; Ezcurra 2016). Similarly, the presence of a vestigial phalanx in digit V comprises another peculiar condition among the members of the clade. Yet, the presence/absence of phalanges in digit V, which is often vestigial, is highly affected by taphonomic biases.
Local palaeofauna
The Varzea do Agudo (¼ Janner) site is one of the most prolific fossiliferous localities from southern Brazil,
from which year-after-year numerous new specimens have been exhumed. This fauna is dominated by traversodontid cynodonts of the genus Exaeretodon (Muller et al. 2020b) . This remarkable abundance has been used to correlate these beds with those from the upper portion of the Ischigualasto Formation ( Langer et al. 2007; Ezcurra et al. 2015; Muller & Garcia 2020; Schultz et al. 2020). As a consequence, the younger Brazilian proterochampsids are divided into two distinct horizons within the Hyperodapedon AZ. Rhadinosuchus gracilis and Cerritosaurus binsfeldi were unearthed from outcrops of the Alemoa complex ( Garcia et al. 2019), which belong to the Hyperodapedon Acme Zone (lower portion; Ezcurra et al. 2015; Schultz et al. 2020), whereas Proterochampsa nodosa comes from Exaeretodon -dominated beds (upper portion; Schultz et al. 2020). In this context, Stenoscelida aurantiacus expands the abundance and diversity of proterochampsids in the Exaeretodon sub-AZ.
The presence of a proterochampsid in the Varzea do Agudo site is not particularly helpful regarding a biostratigraphic approach because proterochampsids occur across the entire Ischigualasto Formation ( Martınez et al. 2013). Nevertheless, the new taxon comprises an additional predatorial taxon for the palaeofauna of this site. The absence of cranial remains precludes an unambiguous interpretation regarding the diet of Stenoscelida aurantiacus , but since all proterochampsids are considered carnivorous animals ( Trotteyn et al. 2013), the null hypothesis is that Stenoscelida aurantiacus exhibited a similar diet. So far, Stenoscelida aurantiacus and Dynamosuchus collisensis comprise the only carnivorous archosauriforms from this site, with the ectetniniid cynodont Trucidocynodon riograndensis ( Oliveira et al. 2010) adding to this context. Other occurrences at the site belong to herbivorous/omnivorous animals: the archosauromorph Hyperodapedon sp. , the traversodontid cynodont Exaeretodon , and the sauropodomorph dinosaurs Pampadromaeus barberenai ( Cabreira et al. 2011) and Bagualosaurus agudoensis ( Pretto et al. 2019) .
The putative mode of life related to water bodies often attributed to proterochampsids are an interesting theme added to the Varzea do Agudo palaeofauna, which previously recorded only fully terrestrial taxa (see above). Currently, the site lacks other organisms that possibly preyed within aquatic environments, such as temnospondyls and phytosaurs. Therefore, if proterochampsids (especially Stenoscelida aurantiacus ) were semi-aquatic predators (following the general morphology of extant crocodilians; Trotteyn et al. 2013), the new taxon expands the ecological diversity within the Varzea do Agudo site. On the other hand, some methods have cast doubts on these habits. Therefore, while some proterochampsids probably had a mode of life related to water bodies, other were more likely terrestrial in habit (e.g. Chanaresuchus bonapartei ; Arcucci et al. 2019; Ezcurra et al. 2021).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.