
Zealeuctra uwharrie, Verdone & Beaty & Holland & Kondratieff, 2019
|
publication ID |
https://doi.org/ 10.5281/zenodo.4758109 |
|
publication LSID |
lsid:zoobank.org:pub:EC8E4749-F916-4F51-82F8-D5C679E06B0E |
|
DOI |
https://doi.org/10.5281/zenodo.4765396 |
|
persistent identifier |
https://treatment.plazi.org/id/038AC112-FFA2-FFAC-0FDF-F915FBCAFE2C |
|
treatment provided by |
Felipe |
|
scientific name |
Zealeuctra uwharrie |
| status |
sp. nov. |
Zealeuctra uwharrie View in CoL sp. n. Verdone, Beaty, Holland & Kondratieff
http://lsid.speciesfile.org/urn:lsid: Plecoptera .speciesfile.org: TaxonName:506050
( Figs. 1–2 View Figs , 4–19 View Figs View Figs View Figs View Figs View Figs )
Material examined. Holotype ♂ U.S.A. — North Carolina: Montgomery Co. , unnamed spring tributary to Little River , SR 1005, Troy Boat Access, N 35.29628, W 79.88583, 16 January 2019, C. Verdone, S. Beaty, V. Holland, B. Kondratieff ( NMNH) GoogleMaps . Paratypes: Montgomery Co., Gold Mine Branch , FS Rd. 555, N 35.41232, W 80.02314, 23 January 2019, C. Verdone, S. Beaty, V. Holland, 6♂, 6♀ ( INHS) GoogleMaps ; Little Creek , SR 1516, N 35.27616, W 79.81779, 16 January 2019, C. Verdone, S. Beaty, V. Holland, B. Kondratieff, 10♂, 6♀ ( CSUIC) GoogleMaps ; Little Rocky Creek , SR 1543, N 35.21038, W 79.93981, 26 January 2019, C. Verdone, D. Fuller 3♂, 1♀ ( INHS) GoogleMaps ; unnamed tributary to Barnes Creek , SR 1303, N 35.44052, W 79.99970, 23 January 2019, C. Verdone, S. Beaty, V. Holland, 17♂, 15♀ ( NCDWR) GoogleMaps ; same location, 2 February 2019, C. Verdone, S. Beaty, V. Holland, 4♂, 6♀ ( WKUC) GoogleMaps ; same location, 12 March 2019, C. Verdone, S. Beaty, V. Holland, 1♀ ( NCDWR) GoogleMaps ; unnamed tributary to Cheek Creek , SR 1563, N 35.24810, W 79.80711, 16 January 2019, C. Verdone, S. Beaty, V. Holland, B. Kondratieff, 2♂ ( CSUIC) GoogleMaps ; unnamed spring tributary to Little River , SR 1005, Troy Boat Access, N 35.29628, W79.88583, C. Verdone, S. Beaty, V. Holland, B. Kondratieff, 15♂, 9♀, 2 larvae, 1 exuvium ( NCDWR) GoogleMaps ; same location, 23 January 2019, C. Verdone, S. Beaty, V. Holland, 4♂, 5♀, 2 larvae, 1 exuvium ( CSUIC) GoogleMaps ; same data, 1♀, 1 exuvium (reared) emerged 13 February 2019 ( NCDWR) GoogleMaps . Randolph Co., unnamed tributary to Uwharrie River , N 35.54490, W 79.97184, 2 February 2019, C. Verdone, S. Beaty, V. Holland, 3♂, 3♀, ( NMNH) GoogleMaps . Stanly Co., unnamed tributary to Mountain Creek , Morrow Mountain State Park, SR 1730, N 35.34196, W 80.11049, 16 January 2019. C. Verdone, S. Beaty, V. Holland, B. Kondratieff, 2♂, 1♀ ( NCDWR) GoogleMaps .
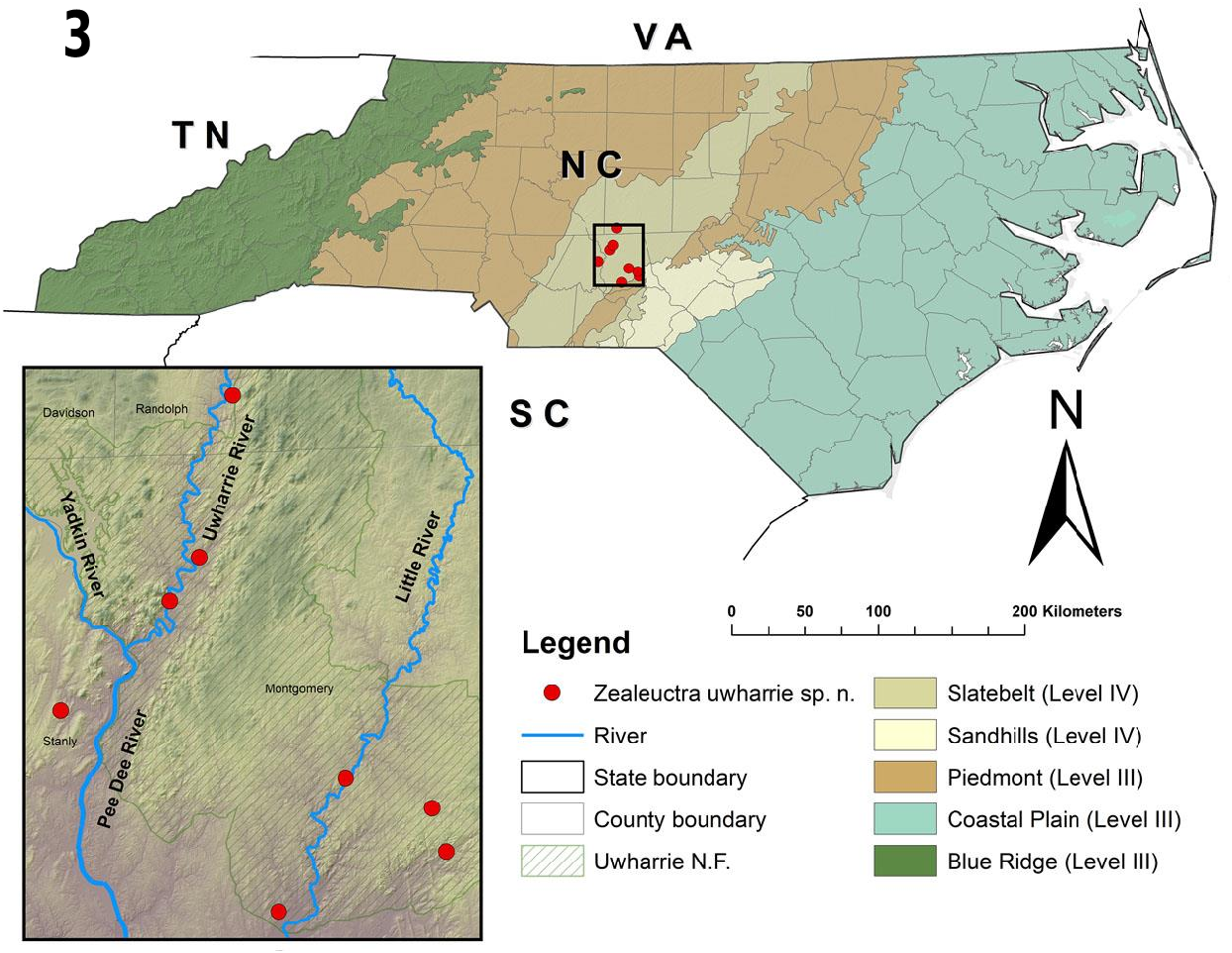
Distribution. U.S.A. – NC ( Fig. 3 View Fig ).
Etymology. The new species is named after the region where the majority of specimens were collected, the Uwharrie National Forest of southcentral North Carolina. The proposed common name is the “Uwharrie Needlefly” ( Stark et al. 2012).
Male. Macropterous. Length of forewings 6.4–7.2 mm (n = 10). Length of body 5.9–6.7 mm (n = 10). General body color dark-brown to black ( Fig. 1 View Figs ).
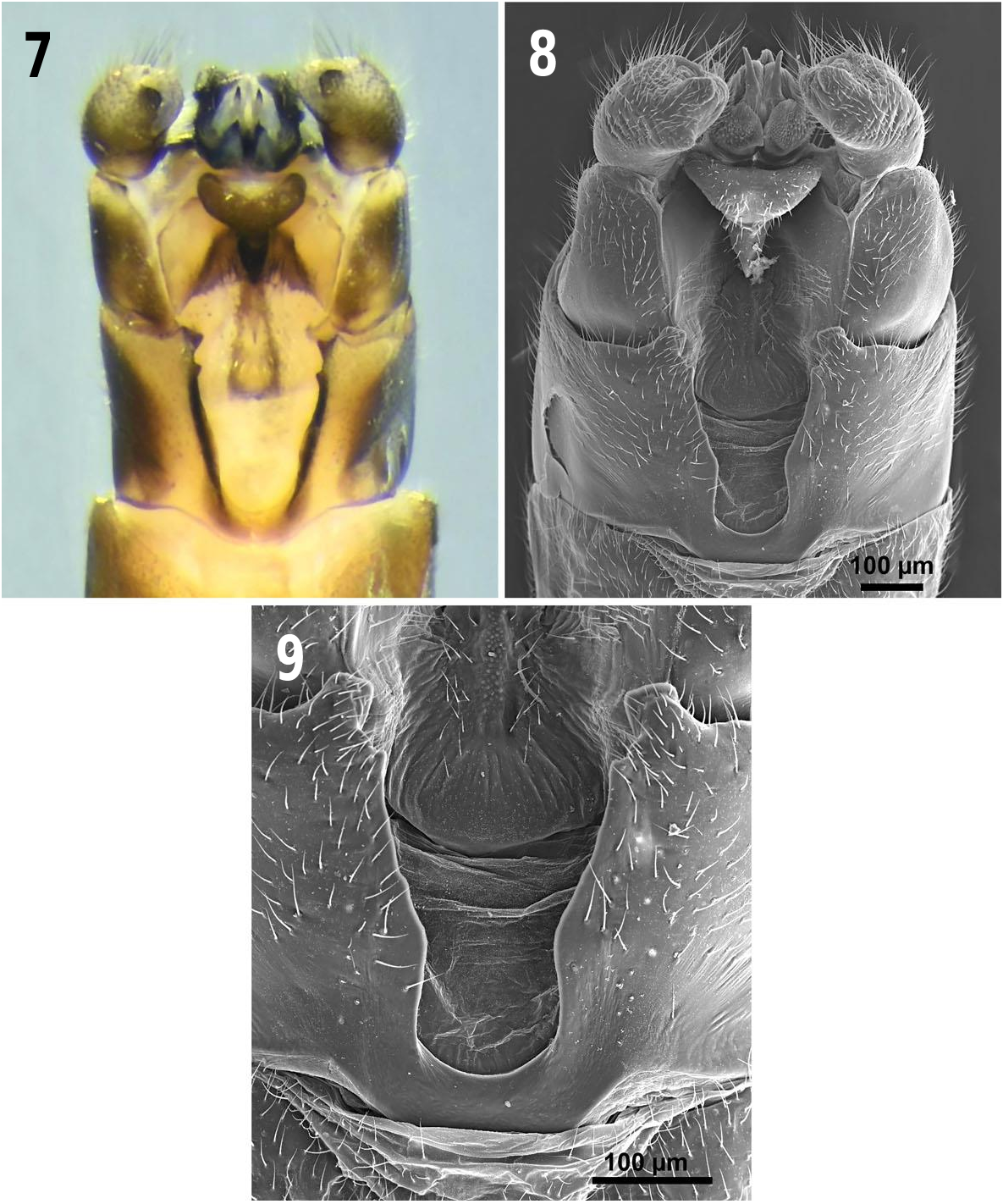
Head: ( Fig. 4 View Figs ). Dorsum of head mostly darkbrown to black; lateral ocelli pale brown; interocellar area often with a slight depression; a piceous, glabrous oval area between lateral ocelli and antennal scape near eye; a pair of small raised humps anterolateral to median ocellus, often slightly darker than surrounding ground color. Antennae uniformly dark-brown; at least 44 segments. Occiput with small, irregular, dark rugosities. Entire head capsule adorned with short, clear clothing hairs, shorter posteriorly.
Thorax: Pronotum ( Fig. 4 View Figs ) subquadrate, slightly longer than wide with anterior pronotal corners rounded; brown overall with darker rugosities; anterior and posterior pronotal furrows darkened lines; median ecdysial line raised and rugose, discal rugosities coalesced in semicircular fashion, resembling a mushroom overall; pronotal sclerotization extending medially beyond posterior margin of the pronotal disc, extensions medially divided by a pale line. Meso- and metathorax heavily sclerotized dorsally and ventrally, with dark brown to black stripe lateral to ecdysial suture. Legs uniformly brown, sometimes slightly darker at the femoraltibial joint; apex of tibiae with 2 stout apical spines. Wings slate grey; venation brown; brown spot present at base of costa ( Fig. 1 View Figs ).
Abdomen: ( Fig. 5 View Figs ). Brown, with a darker brown longitudinal medial sclerotized stripe dorsally, variously developed, and four dark transverse spots on terga 1–8; terga 1–2 mostly pale and membranous; each tergum with a dark brown lateral sclerite, giving the appearance of a lateral abdominal stripe; a dark, thin, linear sclerite in the pleural folds, variously developed on posterior segments; abdominal sterna 1–7 each with two dark spots. In lateral view abdominal sterna heavily setose compared to terga. Vesicle ( Fig. 6 View Figs ) present on posterior margin of sternum 8; length 1.5X width, evenly rounded, entire surface densely clothed in golden setae. Sternum 9 darker than preceding segments, area occluded by vesicle pale and unsclerotized.
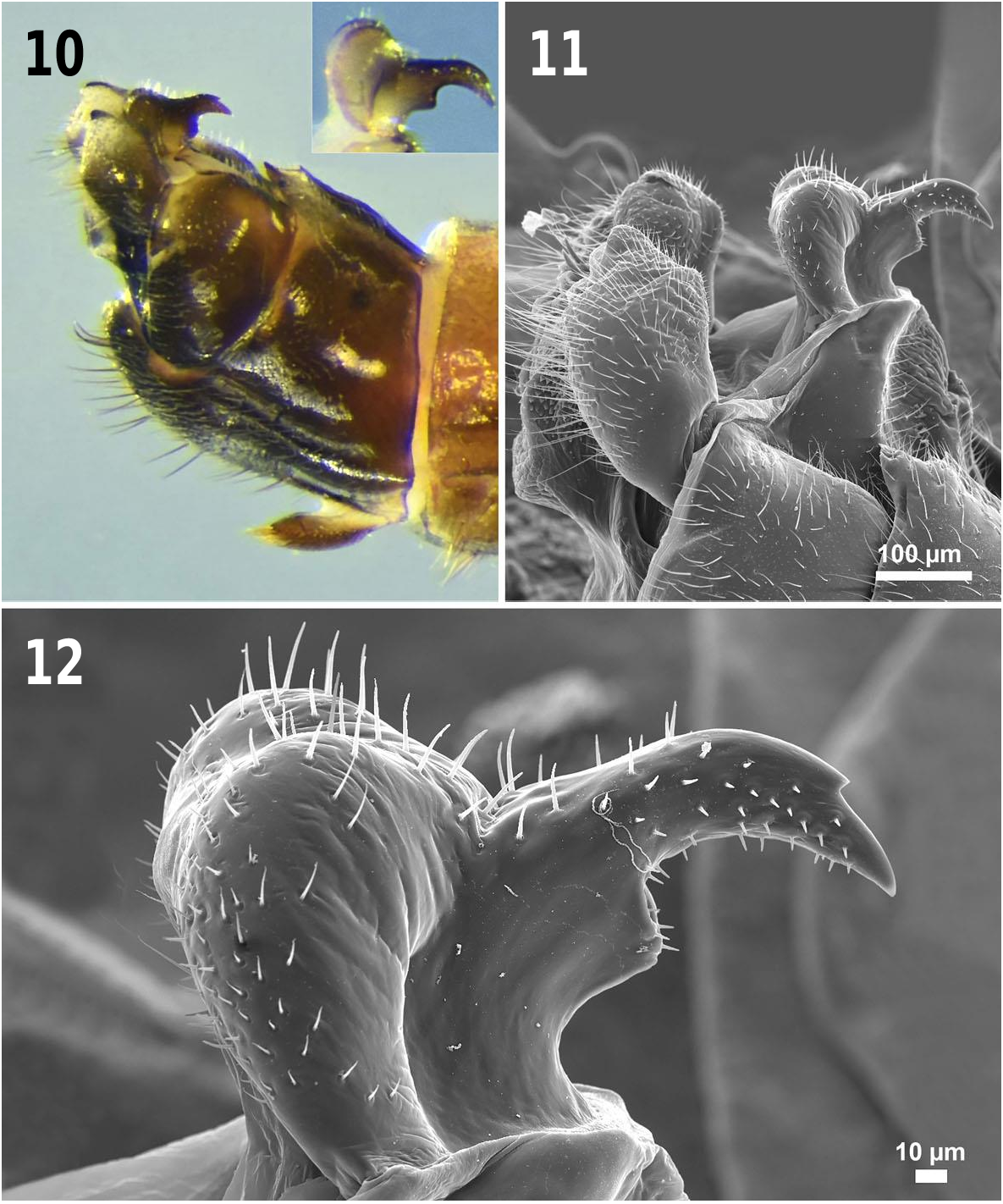
Terminalia: ( Figs. 7–13 View Figs View Figs View Figs ). Tergum 9 deeply cleft, narrowly U-shaped anteriorly, inner margins sinuous, divergent posteriorly ( Figs. 7–9 View Figs ); inner posterolateral margins with small sub-terminal, medially directed points; posterior apices with larger truncated or rounded posteromedially directed projections ( Figs. 8–9 View Figs ). Medial area of tergum nine membranous with a triangular sclerite. Epiproct recurved over tergum 10; base in lateral view expanded, sharply quadrate to broadly rounded ( Figs. 10–12 View Figs ); in lateral view, anterior hooked portion of epiproct, dark brown to black, tapered and strongly curved ventrad with a relatively large dorsal subterminal spine; posterior portion of epiproct adjacent to quadrate base with broad hemispherical lobes that extend caudally ( Figs. 10–12 View Figs ) and laterally in dorsal aspect ( Fig. 13 View Figs ); epiproct with regularly spaced sensilla over much of its surface; dorsoapical areas of the epiproct and subterminal spine, in addition to the lateral areas of the expanded base are glabrous ( Fig. 12 View Figs ). Subanal probe ( Fig. 13 View Figs ) open apically; deeply divided longitudinally for most its length; each side joined together both basally and posteriorly by a membranous sheath bearing dark triangular spinulae apically and posteriorly; dark lateral sclerites envelop the probe laterally and anteriorly; widely separated apically and narrowly basoposteriorly; a pair of long, thin anteromedial sclerites are held internally, becoming membranous subapically, then narrowing to acute well sclerotized points which extend slightly beyond the probe apex ( Figs. 7–8 View Figs ). Cercus one segmented with a small subapical dorsal projection and a darkly sclerotized fingernail-like process on posterodorsal margin; setose ventrally ( Figs. 7–8 View Figs ). Female. Macropterous. Length of forewings 7.6– 7.9 mm (n = 10). Length of body 6.4–8.8 mm (n = 10). General habitus coloration similar to male. Dorsum of abdomen mostly membranous and with a medial sclerotized brown stripe on terga 1– 8 ( Fig. 5 View Figs ).
Terminalia: ( Fig. 14 View Figs ). Sternum 7 strongly produced and heavily sclerotized on posterolateral margins; posteromedial margin lightly sclerotized with a shallow posterior notch and bearing a moderately sclerotized ovoid lobe; sternal lobe length 1.25X width, projecting beyond the posterior margin of tergum 7 recessed in notch for no more than ½ its length; lobe is often in-folded perpendicular to tergum eight. Membranous intersegmental fold posterior to sternal lobe with dark socketed sensillae, often obscured by the in-folding of the posterior margin of sternum eight.
Ovum. ( Fig. 15 View Figs ). Length 164 μm (n = 2); width 137–144 μm (n = 2). General shape oblong; cross section circular; chorionic surface completely smooth; lightly to moderately sclerotized. Despite the ovum’s apparent lack of specialized structures, individual ova readily adhered to the sides of glass vials even when in alcohol.
Mature larva. ( Fig. 2 View Figs ) Length of ♂ body 6.4–7.5 mm (n = 3), ♀ body 9.1 mm (n = 1). General color yellow-brown. Integument covered in fine scales.
Head: ( Fig. 16 View Figs ). Dorsum of head yellow-brown, purplish-brown dorsal reticulations variously developed but delineating ocelli somewhat, markings present lateral to median ocellus; lateral edges of labrum darkly sclerotized. Antennae uniformly yellow-brown, with at least 49 segments.
Maxilla: Lacinia ( Fig. 17 View Figs ) bidentate with two subequal apical teeth; teeth short, truncate and peg-like on pre-emergent specimens; lacinia triangular, evenly receding from base to apical teeth; 11–12 stout marginal setae receding from dorsal apical tooth to the apical ½; marginal setae adjacent to apical tooth short, subsequent seta becoming longer, then shorter; nine stout setae on ventral face below ventral apical tooth; 1–2 prominent hair-like, sub-marginal seta positioned on the basal ½. Galea 1.5X lacinia length or less, broad approximately ⅔ the width of the lacinia; apex bearing 2–3 stout setae. Palpi robust; segments 1–2 subequal, segments 3–5 approximately 2X the length of segments 1–2, segment 5 about 1.3X as long as segment 3 or 4; segments 2–4 bearing 3–6 spines on palpal body.
Mandible: Left mandible ( Fig. 18 View Figs ) with 4 apical teeth; proximal margin with a pectinate comb consisting of 14 long scraping blades (10X as long as wide); basal ⅕ with a tuft of hair- like setae. Right mandible similar to left except pectinate comb consists of 11 teeth, which are 5X as long as wide. Mouthparts of only one specimen were slide mounted, as such the biometrics of other individuals may vary.
Thorax: ( Fig. 16 View Figs ). Pronotum glabrous, margin smoothly contoured laterally; sclerotization extending medially beyond posterior margin of the pronotal disc, medially divided by a pale line. Length of forewing pads 3.75X width; length of hindwing pads 3X width; posterior ⅖ narrowed. Meso- and metabasisterna light brown, glabrous; furcal sternites slightly darker. Legs pale yellow; dorsum of femur with approximately 20 stout setae, mostly in the distal half. Ventral edge of tibia and tarsi with a line of stout setae, ending with 2 stout apical spines dorsal edge of tibia with a sparse fringe of hair-like setae, mostly concentrated on distal half; tarsal segment 3 with 5–6 hair-like setae.
Abdomen: ( Fig. 19 View Figs ). Glabrous, yellow-brown; abdominal segments 1–7 divided by a ventrolateral membrane. Paraprocts fused basally; male paraprocts strongly divergent apically, tapered to triangular points, cleft 9 th tergum visible through integument in pre-emergent males; female paraprocts mostly parallel, subquadrate apically. Cercus with at least 22 segments; each cercal segment with an apical circlet of 12–15 stiff setae, the longest approximately ¾ the length of the respective segment.
Diagnosis. Zealeuctra uwharrie is defined in the males by having an epiproct consisting of a large, quadrate base and a prominent subapical tooth, in addition to the abdomen being marked with a sclerotized dorsal stripe. Females are characterized by having a dorsal abdominal stripe and a moderately sclerotized median lobe that is recessed half its length within the notched posterior margin of the subgenital plate.
Remarks. Adults of Z. uwharrie are superficially similar to Z. fraxina , Z. talladega , and Z. ukayodi . Males of the new species can be distinguished by the details of the epiproct, the cleft of tergum nine, and abdominal sclerotization. The epiproct of the new species tapers to an acute tip with a sub-terminal dorsal spine ( Figs. 10–12 View Figs ). Of the three similar species, only Z. fraxina has a welldeveloped sub-terminal spine. However, the epiproct of the new species has an expanded base which varies from sharply quadrate to broadly rounded in lateral aspect and the anterior portion of epiproct is strongly curved relative to the base ( Figs. 10–12 View Figs ). Whereas, in Z. fraxina , the epiproct base is generally triangular in lateral aspect and the anterior portion is not as strongly curved ( Fig. 20 View Figs ).
The male of the new species is readily distinguished from Z. fraxina by the shape of the cleft, which is narrowly U-shaped anteriorly, divergent posteriorly with the inner margins sinuate, and the posterior apices are truncated or rounded ( Figs. 7–9 View Figs ). In males of Z. fraxina , the cleft is broadly U-shaped, the inner margins are concave, and the posteromedially directed projections on the posterior apices are sharply pointed ( Fig. 21 View Figs , fig. 4 A, Grubbs et al. 2013). Finally, it appears that Z. uwharrie males are the only species of the genus that have a medial sclerotized stripe on the dorsum of the abdomen ( Fig. 5 View Figs ).
Females of the new species are easily separated from Z. fraxina and Z. talladega by the details of sternum seven and abdominal coloration. In the new species, the notch on the posterior margin of sternum seven is shallow and the lobe projects beyond the posterior margin for over half its length ( Fig. 14 View Figs ). Additionally, the lobe on Z. uwharrie is moderately sclerotized. In females of Z. fraxina ( Fig. 22 View Figs ) and Z. talladega , the notch on sternum seven is deep, the lobe is very lightly sclerotized and scarcely projects beyond the posterior margin of sternum seven.
Females of the new species differ from Z. ukayodi in the following: in Z. ukayodi , the lobe on sternum seven is recessed slightly, if at all. Additionally, in Z. ukayodi , sternum seven has a triangular pale area medially that begins near the anterior ¼ and widens posteriorly. In Z. uwharrie , the pale area on sternum seven is restricted to the posterior margin and the lobe is recessed up to ½ its length ( Fig. 14 View Figs ). Finally, the new species has a conspicuous dorsal abdominal stripe ( Fig. 5 View Figs ), which the other species lack.
The larva of Z. uwharrie is only the second species of this genus to be formally described, the other being Z. claasseni ( Frison 1929, Stewart & Stark 2002). Therefore, separation of Z. uwharrie from its congeners is not recommended at this time.
Type locality. ( Fig. 23 View Fig ). The unnamed tributary to the Little River in Montgomery County, North Carolina is a 1 st order stream with a drainage area of 0.22 km 2. The watershed is primarily forest, but the small stream is bordered by several roads, including a gravel landing for a state-maintained boat access on the adjacent Little River. During the initial and subsequent collections in January 2019, we observed one spring source within the stream channel. The water temperature at the spring source was 9.1°C. In-stream habitat consisted of stepped pools containing a mix of boulder, cobble, gravel and silt. Riparian vegetation included American holly ( Ilex opaca Aiton ), American beech ( Fagus grandifolia Ehrh. ) and Christmas fern ( Polystichum acrostichoides (Michx.) Schott ).
Biological notes. Zealeuctra uwharrie is currently known from eight locations in the vicinity of Uwharrie National Forest in the Carolina Slate Belt of North Carolina. The Slate Belt consists predominantly of low water-yielding rocks ( Daniel 1989); consequently, streams in this area are prone to reduced flows and may dry up in the summer months ( Griffith et al. 2002, Beaty et al. 2017). The new species has been documented from 1 st –2 nd order streams with drainage areas ranging from 0.22–6.81 km 2. The elevation range of occurrence localities is approximately 74– 147m. Water temperature taken at four locations at the time of collection ranged from 7.0°–9.4° C. In 2019, adults were active from mid-January to at least mid-March. Larvae were difficult to collect, and an inordinate amount of time was spent looking for the few larvae that were obtained. Like other Zealeuctra ( Snellen & Stewart 1979) , Z. uwharrie may inhabit the hyporheic zone until just prior to emergence. Additional stonefly species often collected in association with Z. uwharrie were Allocapnia wrayi Ross, 1964 and A. rickeri Frison, 1942 , two species commonly collected in the region. Although records of Zealeuctra east of the Appalachians are few, the range extension presented by this new species is perhaps unsurprising considering Zealeuctra are typically found in spring-fed streams that traverse hilly terrain and exhibit seasonally low flows or drying ( Ricker 1952, Snellen & Stewart 1979). Two other recently described Zealeuctra , Z. talladega , and Z. ukayodi also occur in low-elevation southern Appalachian landscapes.






| NMNH |
Smithsonian Institution, National Museum of Natural History |
| INHS |
Illinois Natural History Survey |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |