
Xenolepisma monteithi, Smith, 2015
|
publication ID |
https://doi.org/ 10.3853/j.2201-4349.67.2015.1641 |
|
persistent identifier |
https://treatment.plazi.org/id/03C687D5-6A3D-1237-FEA4-FE18CC2B06AC |
|
treatment provided by |
Carolina |
|
scientific name |
Xenolepisma monteithi |
| status |
sp. nov. |
Xenolepisma monteithi n.sp.
Figs 47–85 View Figures 47–59 View Figures 60–71 View Figures 72–85
Type material. Holotype ♀ (probably not fully mature, HW 0.55) ( QM T228756 on two slides) Queensland: Cape York, Bramwell Station, 12.13866°S 142.62345°E 73m asl, 21.vii.2013, Graeme Smith , pyrethrum spray to bark of Eucalypt GoogleMaps . Paratypes 1 juvenile ♀ ( HW 0.54) ( QM T228757 on two slides) , 1 juvenile ♀ ( HW 0.50) ( QM T 228758 in alcohol with presumed host ants Crematogaster sp.), all same collection data as holotype GoogleMaps .
Diagnosis. This species differs from other described species of the genus most obviously in the presence of macrochaetae on the posterior margins of the nota and in the 6+6 chaetotaxy of the anterior urotergites.
Description
Appearance in life as for all other known members of genus i.e. almost black in colour with distinct 1+1 large white patches on anterior half of mesonotum, transversely across the posterior two thirds of urotergite I and 1+1 patches on anterior margin of urotergite VII (see Smith et al., 2011: plate 2), due to the presence of hyaline scales that allow the colour of the underlying integument to show through; these patches can be very difficult to define in animals that have lost a lot of their scale cover. All appendages appear light brown/white in colour.
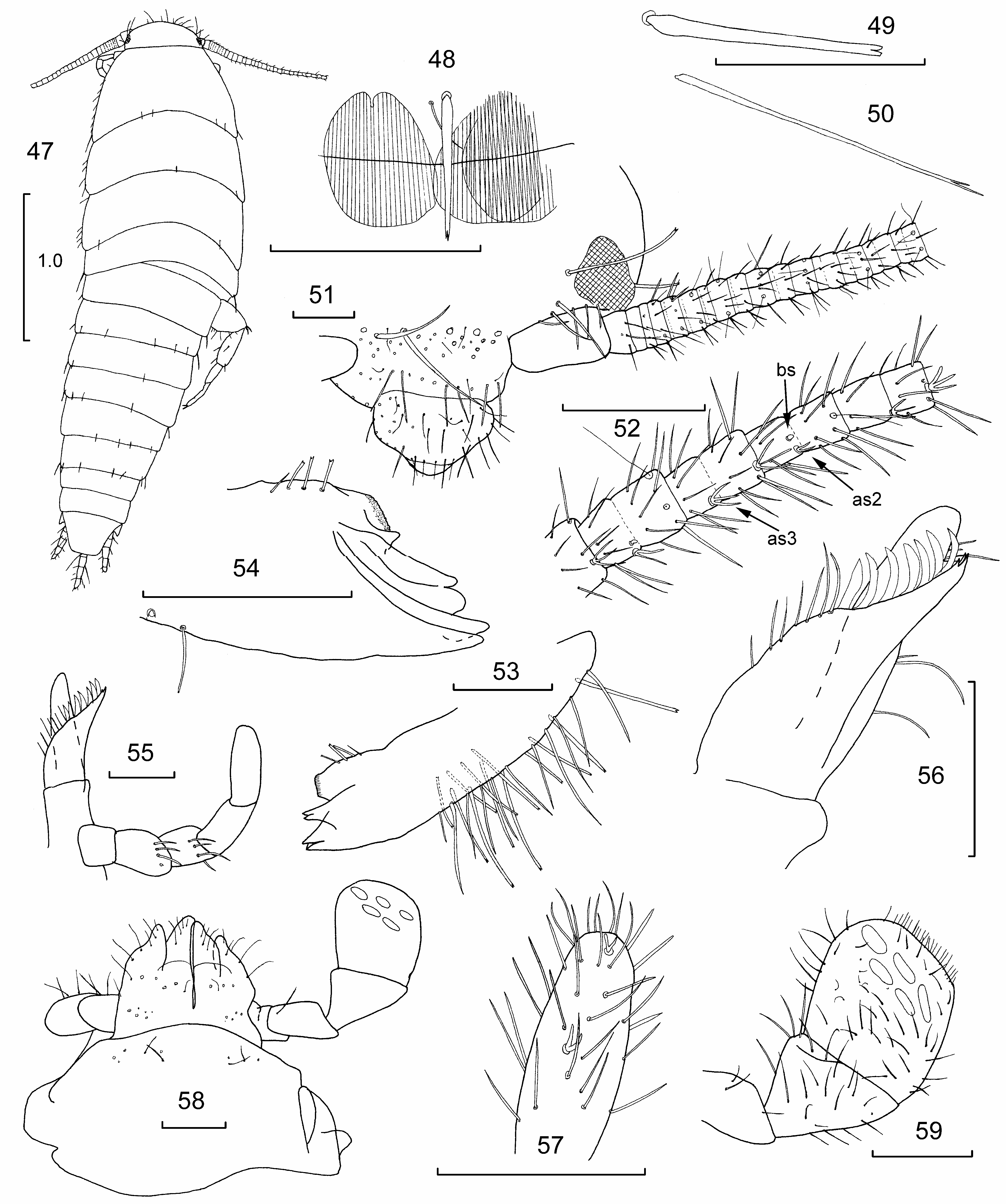
Small insects ( Fig. 47 View Figures 47–59 ), about 3.5 mm long (estimated as abdomen distended in all specimens and probably not fully mature); maximum head width 0.55 mm; antennae incomplete in holotype (>⅓ H+B), probably just a little longer than the thorax; cerci short, less than one tenth H+B; median dorsal appendage incomplete in holotype, slightly longer than cerci exceeding the apex of urotergite X by about the length of this urotergite. Thorax about half the length of the abdomen; abdomen at its base not much narrower than the thorax and not clearly set apart from it.
Integument generally white with blotchy brownish pigment present on the back and sides of the frons, the entire clypeus, labrum and labium including mouthparts although less on the maxillary palp, especially the ultimate article, and absent from around the papillae on the ultimate article of the labial palp. Present on the scape but not pedicel; remainder of flagellum with some pigment in proximal half. Pigment present along sides of nota and also the posterior urotergites. Coxae with intense pigment but remaining articles white (except for hint of pigment along margins of femur), moderately strong pigmentation on prothoracic presternum and thoracic sternites.
Scales with numerous subparallel rays which do not surpass the posterior margin of the scale ( Fig. 48 View Figures 47–59 ), in alcohol mostly dark to lighter brown in colour or hyaline. Scales on top of head but lacking elsewhere on head and absent from mouthparts and antennae, coxa heavily scaled, but scales absent from remaining leg articles. Ovipositor and terminal filaments without scales.
Macrochaetae smooth, some robust and apically bifurcate ( Fig. 49 View Figures 47–59 ), others more slender with delicate apical bifurcations ( Fig. 50 View Figures 47–59 ).
Head ( Fig. 51 View Figures 47–59 ) wider than long; eyes well forward; vertex lacking chaetotaxy; some strong macrochaetae around the eyes, a narrow field of a few strong macrochaetae and several smaller setae between antennae above the clypeus. Clypeus with area of weaker setae subdistally. Labrum with a few setae. —Antennae ( Figs 51, 52 View Figures 47–59 ) with subarticles of most distal surviving articles about as long as wide, with groups of four subarticles forming a chain with repeated sensillae patterns. Specialized asteriform sensillae present in distal two thirds of the flagellum ( Fig. 52 View Figures 47–59 ). Chains with a triramous asteriform sensilla on each of the two most distal subarticles of a chain, a trichobothrium on the following segment and a biramous specialized asteriform sensilla, usually adjacent to a small thin-walled basiconic sensilla (see Adel, 1984) on the basal subarticle of each chain. —Mandibles ( Figs 53, 54 View Figures 47–59 ) with three or four incisor teeth, and a distinct but small molar region and a field of about 20–30 strong, simple or apically bifurcate macrochaetae on external surface in proximal two thirds. —Maxillae ( Figs 55–57 View Figures 47–59 ), lacinia with three small teeth, one shorter than the others, followed by seven lamellate process and seven long setae; maxillary palp short, its distal article subcylindrical about 2.7–3.0 times longer than wide and 1.4–2.0 times longer than the penultimate article, the second article subapically with rosette of somewhat stronger stout, apically bifurcated macrochaetae, third article with similar but weaker rosette of slightly stronger simple setae, last article with fine setae and one or two (more?) biramous asteriform sensillae ( Fig. 57 View Figures 47–59 ). —Labium ( Fig. 58 View Figures 47–59 ), apical article of palp about 0.76–0.81 times longer than wide, with 3+2 large compact sensory papillae apically extending across most of the width of the article ( Fig. 59 View Figures 47–59 ).
Pronotum ( Fig. 60 View Figures 60–71 ) about 1.3–1.4 times longer than meso- and metanota respectively; lacking collar of setae along anterior margin. All nota with short macrochaetae on lateral margins and a submarginal row of 5+6 or 6+6 setae, each seta mostly, but not always associated with a small setula ( Fig. 48 View Figures 47–59 ). Each notum with two closed trichobothrial areas (isolated from margins by scales), those of the prothorax located anteriorly and subposteriorly, while those of the meso- ( Fig. 61 View Figures 60–71 ) and metanota ( Fig. 62 View Figures 60–71 ) are both subposterior. Each trichobothrial area provided with a short trichobothria and a small setula posterolaterally to it ( Figs 63, 64 View Figures 60–71 ).
Presternum ( Fig. 65 View Figures 60–71 ) fairly wide with posterior marginal row of fine delicately apically bifurcate setae. Prothoracic sternum ( Figs 65, 66 View Figures 60–71 ) small, subcordiform, slightly wider at its base than long, with small emargination on each side, several fine marginal setae and 1+1 larger stronger delicately bifurcate submarginal setae, as well as 1+1 fine setae submedially. —Mesosternum ( Fig. 67, 68 View Figures 60–71 ) about 1.2 times as wide as long and not obviously emarginated, with some small marginal setae distally and 1+1 subapical combs of four to five long, apically delicately bifurcate setae; the distance between the combs being slightly more than the length of each comb. —Metasternum ( Figs 69, 70 View Figures 60–71 ) similar in size and chaetotaxy to mesosternum.
Legs ( Figs 65, 67, 71 View Figures 60–71 ), tibia L/W PI 2.1–2.5, PII 2.5–3.1, PIII 3.1–3.4; tarsi L/W PI 5.4–6.0, PII 5.8–6.8, PIII 5.8–7.3; sclerite immediately basal to each coxa of PI with two long stronger, apically bifurcate setae. —Coxae of all legs scaled with strong apically bifurcate macrochaetae, some robust and other more delicate, and finer smaller setae along the outer margins; ventral surface medially with a comb of 11–12 simple short setae and on dorsal surface anteriorly combs of two or three delicately apically bifurcate macrochaetae near the outer margin and medially two long fine setae posteriorly. —Trochanter fairly large with setae only. —Femur with one longer stout and two smaller macrochaetae at medial posterior angle and some stronger setae along the ventral ridge distad of the medial angle, rest of ventral surface covered in fine setae. —Tibia with two stout macrochaetae distally, one of which is the same length as the apical spur; ventral face with one stout macrochaeta about one third the distance along the tibia, another stout macrochaeta on the posterior margin and another on the anterior margin near the distal end; rest of surface covered in fine setae. —Tarsi consisting of three articles covered with setae. —Pretarsus with two lateral claws and a medial slightly shorter empodial claw.
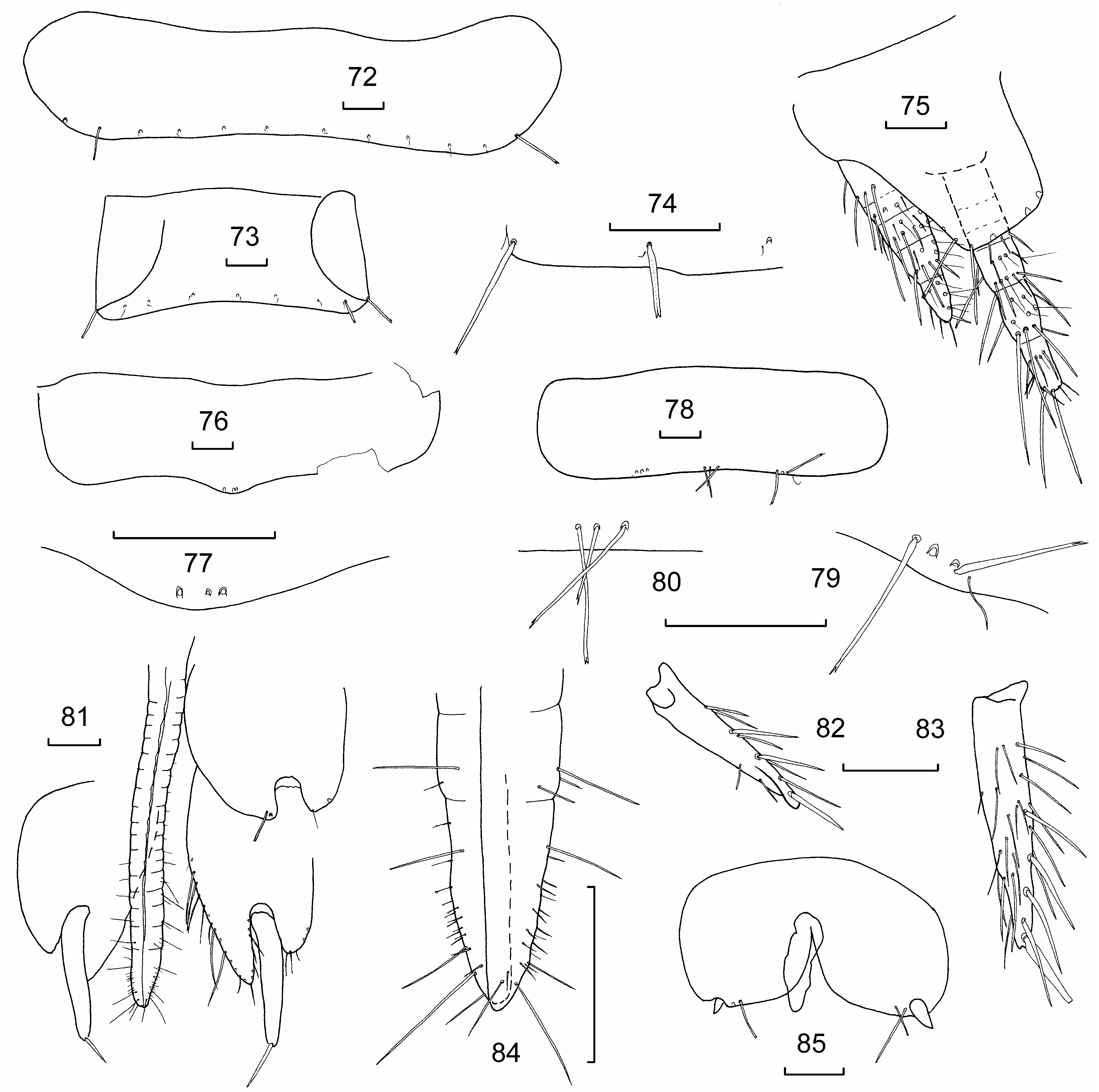
Abdominal chaetotaxy summarized in Table 2. Urotergites I–IX ( Figs 72, 73 View Figures 72–85 ) with 1+1 infralateral macrochaetae almost ½ the length of the corresponding tergite located on the margin of the tergite. Urotergites I–V with additional 5+5 distinctly submarginal setae which are shorter than the infralateral macrochaetae ( Fig. 74 View Figures 72–85 ) and somewhat superficially similar in appearance to the abiesiform setae of Australiatelura spp. (see Smith & McRae, 2014: 106) distributed with almost equal spacing anterior to the posterior margin, the number of macrochaetae decreasing progressively to only 3+ 3 in total on IX. Most urotergal setae associated with a small cilia located between the insertion of the seta and the margin and usually slightly laterad to it ( Fig. 74 View Figures 72–85 ). Urotergite X ( Fig. 75 View Figures 72–85 ) trapezoidal, with 2+2 macrochaetae along the almost straight posterior border (more concave in juvenile paratype) and at least one macrochaeta on lateral border.
Urosternite I without combs. Urosternite II with a narrow medial comb of three long, thin apically bifurcated macrochaetae ( Figs 76, 77 View Figures 72–85 ). Urosternites III–VII ( Fig. 78 View Figures 72–85 ) with 1+1 sublateral combs ( Fig. 79 View Figures 72–85 ) each of 1–4 delicately apically bifurcate macrochaetae associated with a small cilia and in one case a setula between two of the macrochaetae of one comb and one medial comb ( Fig. 80 View Figures 72–85 ) of 2–4 long, thin macrochaetae. The lateral combs on III more mediad than those of the more posterior segments. Coxites VIII of ♀ with 1+1 combs of two macrochaetae internal to stylets ( Fig. 81 View Figures 72–85 ). Internal process of coxite IX of ♀ ( Fig. 81 View Figures 72–85 ) about 1.8 times as long as broad at the base and 1.6 times longer than the external process. —Stylets on urosternites VIII and IX in ♀ holotype ( Figs 82, 83 View Figures 72–85 ). —Ovipositor ( Figs 81 and 84 View Figures 72–85 ) only just surpassing apex of internal processes of urosternite IX; with about 18 articles and only fine setae. It is however quite likely that the ovipositor of the holotype has not achieved its maximum length as the juvenile female paratype with a head width only slightly less than the holotype has a very rudimentary ovipositor and stylets only just appearing on coxites VIII ( Fig. 85 View Figures 72–85 ).
Terminal filaments short, cerci consisting of only about five articles and median filament of about seven, the latter only extending beyond the posterior margin of urotergite X by about the length of the urotergite ( Fig. 75 View Figures 72–85 ); terminal filaments with numerous small trichobothria and some small and also strong macrochaetae.
Mature males unknown. Etymology. The species is named for Dr Geoff Monteith as a way of thanking him for introducing me to his pyrethrum spray sampling method (outlined in Baehr, 1995). The method has proven remarkably effective at non-destructively sampling complex habitats and has opened my eyes to the importance of the bark of standing trees as a habitat for so many Australian lepismatid species.
Remarks
Xenolepisma monteithi is the firstAustralian native species of the subfamily Lepismatinae to be described. This subfamily has its greatest number of species in the Afrotropical and Palearctic regions. The only other species of the subfamily reported from Australia is the cosmopolitan anthropophilic European household silverfish Lepisma saccharina Linnaeus, 1775 which was recorded doubtfully by Silvestri (1908) and reported as common within storage facilities in the “larger towns of the Commonwealth” by Womersley (1939). The genus Xenolepisma is also known from southern Africa, India and peninsula Malaysia. Smith et al. (2011) discussed the history of this genus when describing Xenolepisma penangi and also presented a key to the species of the genus. The new species would key to X. subnigrina Silvestri at the first couplet but can easily be separated from it by the presence of macrochaetae on the posterior margins of the nota (absent in all other species described from the genus) and the 6+6 macrochaetae on the posterior margin of the more anterior urotergites. Smith & Watson (1991) noted the presence of Xenolepisma in Australia based on a single specimen collected at Bundeena near Sydney in New South Wales (ANIC), specimens collected near Townsville (ANIC) and with termites near Coen (ANIC), both in Queensland ( Fig. 1 View Figure 1 ). There are also two records from the Northern Territory with a specimen collected from a mangrove stump near Darwin (ANIC) and Conway (1992) reported Xenolepisma sp. from a dormant nest of the Red Honey Ant ( Melophorus bagoti Lubbock ) at Alice Springs. Presumably the genus is widespread but not commonly encountered.
ACKNOWLEDGMENTS. For the decision to erect a new genus for Qantelsella I am very grateful for the advice of Dr Luis Mendes, (Lisbon, Portugal) and Dr Rafael Molero-Baltanás (Cordoba, Spain) both of whom discouraged me from my original plan to include the species within Ornatilepisma . They willingly shared their extensive experience, including of the African fauna, resulting in this revision of my earlier draft. I would also like to thank Dr Geoff Monteith, formerly QM, for collecting advice, the Queensland Department of Environment and Heritage Protection for permission to collect within the Bladensburg National Park, Mr Derek Smith of the Australian Museum for identification of the ant species from Bramwell Station and Dr Shane McEvey, also of the Australian Museum, for assistance with the illustrations.


| QM |
Queensland Museum |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |