
Sympterichthys moultoni, Last & Gledhill Csiro, 2009
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2252.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/E94B87D0-FFD0-FFAA-7CD9-C327BFC60904 |
|
treatment provided by |
Felipe |
|
scientific name |
Sympterichthys moultoni |
| status |
sp. nov. |
Sympterichthys moultoni View in CoL sp. nov.
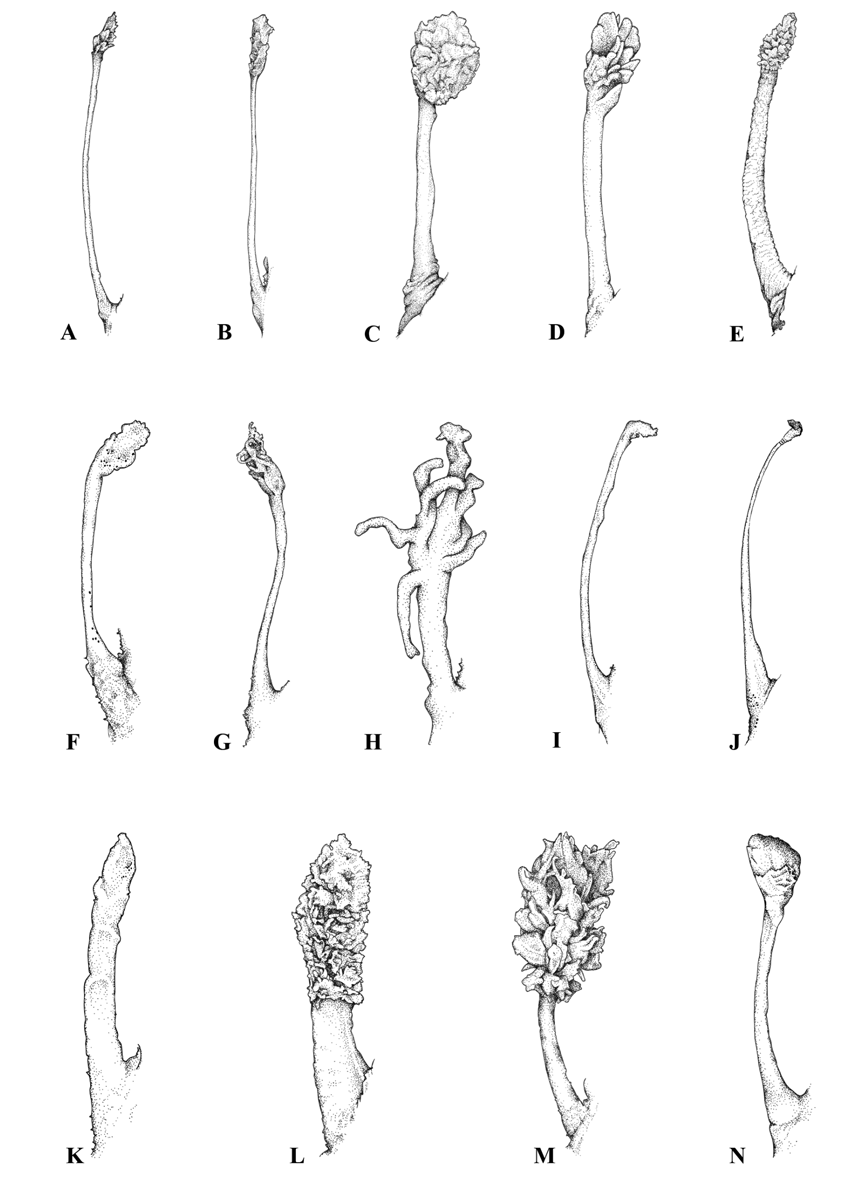
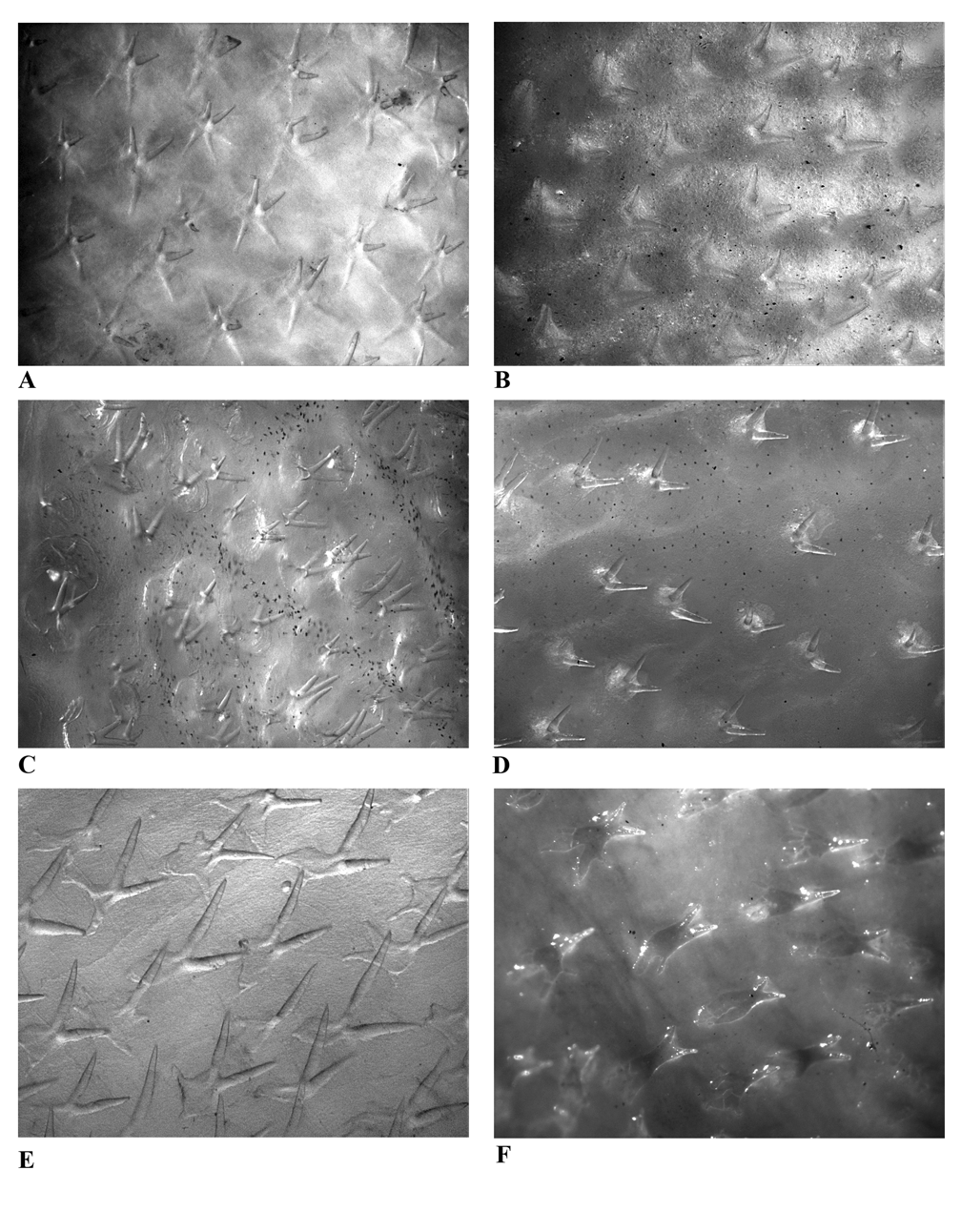
Figs 2 View FIGURE 2 , 4 View FIGURE 4 , 6 View FIGURE 6 , 20, 21; Tables 5, 7–10
Holotype: CSIRO H 3773–03 View Materials , 45.1 mm SL, east of Cape Howe , New South Wales, 37° 25'S, 150° 16'E, 161– 184 m, 1 Sep. 1994. GoogleMaps
Paratypes: 3 specimens (30–39 mm SL): CSIRO H 3792–02 View Materials , 29.5 mm SL, south-east of Lakes Entrance, eastern Bass Strait , Victoria, 38° 36'S, 148° 32'E, 125 m, 27, Aug. 1994 GoogleMaps ; CSIRO H 4461–01 View Materials , 39.3 mm SL, south of Gabo Island , Victoria, 37° 45'S, 150° 03'E, 133–136 m, 7 Dec. 1996 GoogleMaps ; NMV A 29407–001 View Materials , 36.1 mm SL, south-east of Lakes Entrance, eastern Bass Strait , Victoria, 38° 58'S, 148° 33'E, 211 m, 5 Mar. 1984 GoogleMaps .
Additional material: H 6815–01, 38.9 mm SL, south-east of Cape Barren Island, Bass Strait , Tasmania, 40° 33'S, 148° 49'E, 105–150 m, 13 May 2003 GoogleMaps .
Diagnosis. Member of the genus Sympterichthys with a combination of the following characters: esca large, size variable, about 56% of illicium length; illicium thick, fleshy, with some dermal spinules on basal half, length 16–21% SL, 2.1–3 times in head length; esca similar thickness to rest of structure; body entirely covered with close-set, non-imbricate scales; scale bases rudimentary, margins irregular, with long bifurcate spinules (except for those closely associated with pores of the acoustico-lateralis system); spinules mainly adpressed, originating from posterior region of scale base, apical spines moderately divergent; second dorsalfin rays 15–16, fin base 61–66% SL; length of second dorsal-fin spine 0.8–0.9 times length of longest ray of second dorsal fin; 7–8 anal-fin rays; 7–8 pectoral-fin rays; upper body uniformly pale pink; membranes between distal portions of rays of dorsal, caudal and pelvic fins dark.
Description. D1 2 (2, n= 3 paratypes); D2 15 (15–16); A 7 (7–8); Pc 8 (7–8); Pv i, 4 (i, 4); C 1 (1) + 6 (6) + 2 (2) = 9 (9); Vt 10 (9–10) + 11 (11–12) = 21 (21).
Body short to moderately elongate, moderate to strongly compressed; upper anterior profile strongly convex, upright before second dorsal fin; upper margin of eye well below dorsal margin of head, at level of illicial base; head almost oval when viewed anteriorly; nape humped very slightly; anterior ventral profile strongly convex; abdomen expanded slightly; caudal peduncle short, length 6.2 (4.2–5.2)% SL. Head length 50 (45–55)% SL; snout variable in length, 4.8 (5.6–6.6) times in head; eye very small, lateral, somewhat embedded, 7.4 (7.4–9.6) times in head length; gill opening small, aperture subequal to pupil, located posterodorsal to insertion of pectoral fin; nostrils small, distinct, surrounded with spinules in holotype; posterior opening almost dorsal to anterior opening, separated slightly from orbit. Mouth very small, narrow, terminal to slightly inferior, weakly protractile; upper jaw oblique, 3.4 (2.8–3.5) in head; lips fleshy; angle of jaw deeply recessed into groove, situated below middle of eye; tongue narrowly rounded apically. Teeth (in paratype CSIRO H 4461–01) elongate, caniniform, similar in shape in both jaws and few in number; in single row along inner margin of maxilla, in 1–2 rows in lower jaw; vomer apparently edentate (mouth not dissected).
Skin very thick, uniformly covered with small-based, spinulose scales with elongate bifid apical spines; no separate wart-like patches of skin; no obvious dermal flap present on mid-arm of pectoral-fin; dermal flaps absent from body. Body scales close-set, distributed over entire body and over basal half of dorsal and anal fins; spinules on body more or less equidistant from each other, aligned in ill-defined diagonal rows or irregularly; scale bases deeply embedded, not within raised mound of thickened skin. Spinules elongate, mostly embedded, bifurcating within skin, only tips of apical spines exposed; apical spines strongly divergent, not connected by integument, mainly adpressed, orientation variable but frequently posteroventral; those on belly not significantly smaller than those above; scale bases variable in shape, plate-like, with strongly irregular margins, their width subequal to or smaller than spinule length; scale base extremities blunt; spinules arising obliquely from posterior margin of scale base. Basal halves of both dorsal fins (including membranes), anal fin, pelvic fin, and pectoral fin (including all of arm) covered with spinulose scales; distal halves of these fins naked, no scales arranged in series along lateral margins of fin rays; spinulose scales confined to base of caudal fin, holotype with 2 more posteriorly placed sensory scales, otherwise fin naked. Illicium base weakly spinulose, main stem with a few scales on basal half; skin covering stem thick, forming a wide, pouch-like covering around much thinner ossified ray. Scales of acoustico-lateralis system bicuspid, horseshoe-shaped, well demarcated from main body spinules; spine apices overlain with narrow or broad, lobe-like integument; sensory scales variably separated, arranged in obvious series on head and along tail, spinulose scales largely absent from areas connecting these scales.
*Distance from the base of the third dorsal-fin spine to the origin of the second dorsal fin.
Illicium terminal on snout, asparagus-shaped; slender, relatively short, 3.0 (2.1–2.7) times in head, 1.2 (0.8–1.1) times in length of second dorsal-fin spine; apex of esca just short of base of third dorsal-fin spine when fin depressed; partly retractable into shallow groove on either side of first dorsal fin; esca moderately enlarged, finely filamentous, about 3 times in length of illicium, emanating from a equally broad, tubular stem; illicial base not bulbous. First dorsal fin reduced, short-based; spines close together, second dorsal-fin spine almost confluent with base of illicium, slightly longer than third spine; fin membrane thickened, fleshy, expanded almost to apex of each spine (third spine visible distally); posterior extension of membrane very short, terminating forward or below apex of erect third spine; anterior elements of both fins weakly recurved; first dorsal-fin base 3.3 (2.7–3.1) times in second dorsal-fin base. Second dorsal fin low, barely incised, anterior and penultimate posterior rays longest; rays simple; fin base elongate, 66 (61–65) % SL; longest ray of second dorsal fin 0.9 (0.8–0.9) times in longest dorsal-fin spine; basal membrane relatively thick, concealing bases of all fin rays, covering about two-thirds of anterior fin rays and about half of posterior rays. Anal fin short, base fleshy, margin weakly incised; penultimate posterior rays longest; anal-fin base 1.8 (1.9– 2.2) times in second dorsal-fin base. Pectoral fin short, weakly arm-like, elbow extending well beyond gill opening; fin rays digitiform, slender, membranes deeply incised, not increasingly so posteriorly, tips flexible, filamentous. Pelvic fin short, narrow-based; rays slender, deeply incised; anterior spine short, embedded and indistinct; fin located on ventral surface, directed ventrally, base aligned horizontally; interpelvic space narrow, usually convex. Caudal fin small, narrowly rounded; length 3.9 (2.9–3.5) times caudal peduncle depth.
FIGURE 20. A. Collection localities for Sympterichthys moultoni sp. nov., holotype () and paratypes (•); and B. likely type locality of S. unipennis () , southeastern Tasmania. Locations are approximate and may represent multiple records.
Coloration. In life: Body uniform pale pinkish (becoming white in preservative), no evidence of spots or other markings on any of the types. In holotype, fins pale; fin membranes on distal half of caudal and pectoral fins dark, other membranes translucent.
In preserved paratypes: darker brownish areas more extensive; membranes of first dorsal fin, pelvic and pectoral fins, anterior part of second dorsal fin, and posterior parts of second dorsal and anal fins pigmented; smallest specimen (CSIRO H 3792–02) with more uniformly pigmented caudal fins.
Size. Probably a small species, to at least 45.1 mm SL (ca. 61 mm TL).
Distribution. Known from outer continental shelf of eastern Australia from Cape Howe, New South Wales (37° 25'S, 150° 16'E), to eastern Bass Strait, Victoria (38° 58'S, 148° 33'E) in 125–211 m, possibly shallower. An additional specimen collected off Cape Barren Island (Tasmania) was taken in 105– 150 m.
Etymology. Named in honour of the efforts of Peter Moulton, a fisheries biologist with the former Victorian Institute of Marine Science (now the Marine and Freshwater Resources Institute), whose interest in temperate Australian fishes led to the collection of the first specimen of this species, as well as many specimens of hitherto poorly known handfishes. Proposed vernacular name: Moulton’s Handfish.
Comparisons. Sympterichthys moultoni superficially resembles P. compressus in body shape and in having a uniform pale pink coloration (fading to white in preservation). However, these species differ markedly in squamation and characters defining the genera. Sympterichthys moultoni is covered in prostrate, bifid spinules originating at the posterior end of plate-like, scale bases (rather than erect bifid spinules located centrally on stellate scales), and it has a thick illicial sheath (its thickness similar to the esca rather than having a very slender illicium without an obvious sheath), and almost smooth fin rays (rather than being densely covered with spinules).
Sympterichthys moultoni (ca. 30–45 mm SL) differs from the only known specimen of its congener S. unipennis (44 mm SL) in more than 60% of morphometric characters and ratios evaluated. These include having: a deeper (depth at anal-fin origin 21–24 vs. 20% SL; depth at second dorsal-fin origin 41–47 vs. 37% SL) and wider body (maximum width 19–29 vs. 13% SL, although the S. unipennis holotype may have been flattened slightly during preservation); a longer pelvic-fin (length 20–24 vs. 17% SL), snout (length 7.7–11 vs. 7.0% SL), maxilla (length 15–16 vs. 14% SL), longest ray of second dorsal-fin (length 21–26 vs. 19% SL), caudal peduncle (length 4.2–6.2 vs. 1.9% SL), and caudal fin (length 34–36 vs. 32% SL); a wider interorbit (width 9.6–13 vs. 8.2% SL); and a shorter anal-fin base (length 29–33 vs. 36% SL), and second dorsal-fin rays (length 15–17 vs. 22% SL). It also differs in a number of ratios, including: length of longest dorsal-fin spine 0.8–0.9 vs. 1.3 times length of longest ray of second dorsal fin, and length of second dorsal-fin base 1.9–2.2 vs. 1.8 times length of anal-fin base.
Remarks. All five known specimens of Sympterichthys moultoni have been collected since the 1980s using benthic sled or demersal trawl, in about 100– 200 m. Like all handfishes occurring off southeastern Australia, populations of this species may have been impacted by commercial fishing in the region. Little is known of its life history or preferred habitat.
An additional specimen (CSIRO H 6815–01), which was collected recently by D. Maynard off Cape Barren Island (Bass Strait), was tentatively identified as Sympterichthys moultoni . It was excluded from the type series as it differs markedly in the following morphometric ratios: a deeper tail (depth at anal-fin origin 26 vs. 21–24 % SL); wider body (maximum body width 31 vs. 19–29 % SL); a longer pre-second dorsal-fin distance (length 38 vs. 36–37 % SL), anal-fin base (length 35 vs. 29–33 % SL), pectoral-fin (length 26 vs. 21– 25 % SL), third dorsal-fin spine (length 22 vs. 15–17 % SL) and caudal fin (length 50 vs. 34–36 % SL); a larger eye (horizontal diameter 8.6 vs. 5.8–6.8 % SL); a longer interdorsal space (distance between bases of third dorsal-fin spine and second dorsal fin 31 vs. 24–28 % SL); a more slender body anteriorly (depth at origin of second dorsal fin 39 vs. 41–47 % SL); and a shorter caudal peduncle (length 2.9 vs. 4.2–6.2 % SL) and pre-illicial distance (length 3.8 vs. 4.5–5.7 % SL). It also differs in the following ratios: head length 3.1 vs. 2.2–3.1 times illicium length, length of second dorsal-fin spine 1.3 vs. 0.8–1.2 times illicium length, caudal-fin length 4.7 vs. 2.9–3.9 times caudal-peduncle depth, head length 6.0 vs. 7.4–9.6 times eye length and illicium length 32 vs. 33–47% of head length. Otherwise, this specimen is superficially similar to S. moultoni , but without additional material we cannot determine if differences in morphometric characters reflect intraspecific or interspecific variation.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.