
Pyura beta, Skinner & Rocha & Counts, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4545.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:276F1D88-3186-4D14-B0BF-F345D043382B |
|
DOI |
https://doi.org/10.5281/zenodo.5944127 |
|
persistent identifier |
https://treatment.plazi.org/id/03ED87D1-A254-B349-FF24-F8E7E072FE5D |
|
treatment provided by |
Plazi |
|
scientific name |
Pyura beta |
| status |
sp. nov. |
Pyura beta View in CoL sp. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Material examined: Holotype : MZUSP552 View Materials Isla Solarte, mangrove roots, R. M. Rocha, 7/VI/2009 Paratypes: MZUSP 0 0 0 41, Baleeiro south point, São Sebastião, SP, 23/V/1961, Calypso Expedition; DZUP—PYU-022
Cabras Island, São Sebastião, SP, 16/II/1997, 1 ind., R.M. Rocha; DZUP PYU-53 , Oratório Bay , Alcatrazes Island, SP, 2/V/2002 , 1 ind., R.M. Rocha; DZUP PYU-100 , Crawl Key , Bocas del Toro, R. M. Rocha, 25/VIII/2006 ; DZUP PYU-99 , Bastimentos , Bocas del Toro, R. M. Rocha, 4/VIII/2008 ; DZUP PYU-102 Isla Solarte , Bocas del Toro, R. M. Rocha, 7/ VI/2009; DZUP PYU-101 , Isla Solarte , Bocas del Toro, R. M. Rocha, 11/ VI/2009; DZUP PYU-104 , Solarte , Bocas del Toro, R. M. Rocha, 10/ VI/2011; DZUP PYU-75 , Escalvada Island , Guarapari, ES, 27/I/2012 , 1 ind., G.A. Gamba; MZUSP557 View Materials , Amarração Island , Angra dos Reis, 13/III/2014 , 1 ind., L.F.Skinner; CZFFP-ASC38 Sardinhas Bay , Angra dos Reis, 13/III/2014 , 1 ind., L.F. Skinner; CZFFP-ASC39 Comprida Island , Cabo Frio, RJ, 6/V/2014 , 1 ind., L.F. Skinner; DZUP PYU-103 , Solarte , Bocas del Toro, R. M. Rocha, 19/ VI/2014.
Additional material: DZUP PYU-95 Vitória Island , São Sebastião, SP, 8/IX/1998 , 1 ind., R.M. Rocha; DZUP PYU-57 , Montão de Trigo Island , São Sebastião, SP, 1/V/2002 , 1 ind., 10-12m, R.M. Rocha; DZUP PYU-138 , Bastimentos , Bocas del Toro, R. M. Rocha, 4/VIII/2008 ; DZUP PYU-137 , STRI Bay , Bocas del Toro, R. M. Rocha, 5/IX/2008 ; DZUP PYU-66 and Pyu-64, Sumítica Island , São Sebastião, SP, 2/XII/2008 , 1 ind., L.P. Kremer; DZUP PYU-63 , Ponta Grossa bay , Ilha Bela, SP, 03/XII/2008 , 1 ind., L.P. Kremer; DZUP PYU-65 , Serraria Island , São Sebastião, SP; 4/XII/2008 , 1 ind., L.P. Kremer; DZUP PYU-130 , Bastimentos , Bocas del Toro, R. M. Rocha, 27/I/2009 ; DZUP PYU-96 , Escalvada Island , Guarapari, ES, 27/I/2012 , 1 ind., R.M. Rocha; CZFFP- ASC42 Parnaioca bay , Grande Island, Angra do Reis, RJ, 4/ VI/2013, 1 ind., L.F. Skinner; CZFFP-ASC40 Lopes Mendes beach, Grande Island, Angra dos Reis, RJ, 5/ VI/2013, 5 ind., L.F. Skinner; CZFFP-ASC41 Jorge Grego Island , Angra dos Reis, RJ, 6/ VI/2013, 1ind., L.F. Skinner; DZUP PYU-98 , Palmeiras Point , Grande Island, RJ, 10/ IX/2013 , 1 ind, L.F. Skinner; DZUP PYU-97 , Jorge Grego Island , Angra dos Reis, RJ, 11/IX/2013 , 2 ind., L.F. Skinner; CZFFP-ASC43 Dois Rios bay , Grande Island, Angra dos Reis, RJ, 12/IX/2013 , 2 ind., L.F. Skinner; DZUP PYU-94 Papagaios Island , Cabo Frio, RJ, 09/V/2014 , 2 ind., L.F. Skinner.
Etymology. The species name is derived from the coloration of the tunic in resemblance to that of the common beet, Beta vulgaris.
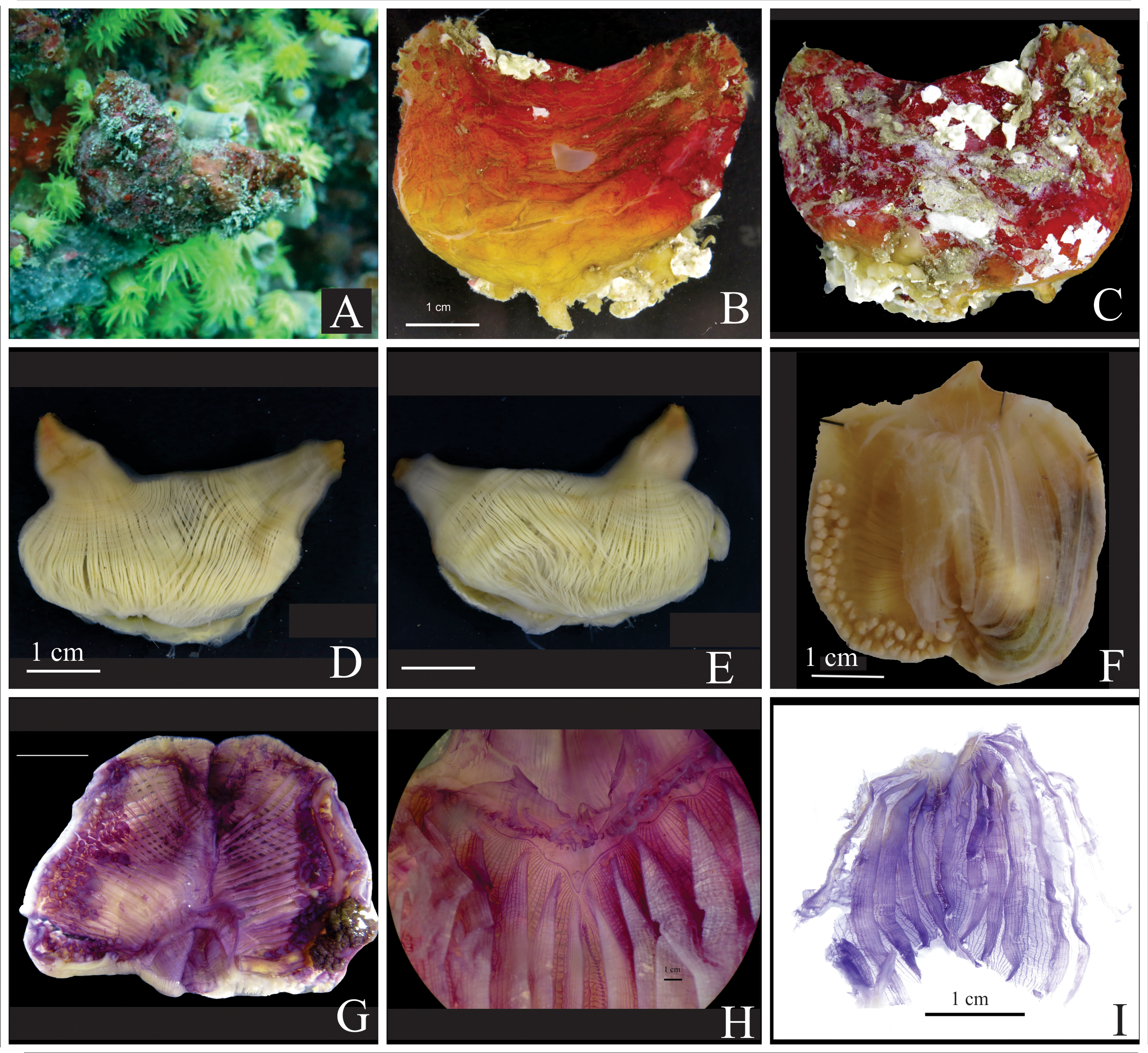
Description. Living specimens present dark red or wine coloration often resembling the coloration of a beet ( Fig. 1A, B, C View FIGURE 1 ), sometimes with a tinge of yellow towards the base of the animal. It loses some coloration after a long fixation. The tunic is hard, 2–3 mm thick, with divisions forming small patches on the surface ( Fig. 1D View FIGURE 1 ), and frequently, but not always, masked by epibionts and debris. The animal is elongated and attaches along the ventral margin to mangrove prop roots, to exposed coral skeleton or rocky walls in shallow waters (less than 5 m in Panama but to 15 m in Brazil), in a vertical position, with the oral siphon apical and the atrial siphon short and posterior. The siphons are surrounded by tubercles ( Fig. 1C View FIGURE 1 ). The holotype is 11 cm in total length, but individuals from 7 to 8 cm long are more common and this was the maximum size found in Brazil. The color of the inner tunic lining depends on the duration of fixation. Freshly fixed specimens present red or wine coloration; however, over the course of time, the tunic shows a gradient from red to white.
With the tunic removed, the holotype is 8.5 cm at the longest length; the oral siphon is 2 cm long and the atrial siphon is 1.5 cm long. Body wall musculature is robust. Thick longitudinal bands composed of many muscle fibers radiate from the siphons and terminate at the level of the gonadal and intestinal region, where those bands divide into numerous thinner fibers leaving this area more transparent ( Fig. 2A, B, D View FIGURE 2 ). The number of muscle bands varies; the right side of the body often has 21–24 bands, and the left side 23–25. Circular musculature around the siphons is also very strong and forms wide bands at the base of both siphons covering the longitudinal bands. The siphons present a variation of red and yellow in recently preserved animals. Internally, the siphons have a velum, but spinules are absent.
Oral tentacles are uncolored, laterally flattened, and triangular with a very wide base. First order ramifications are arranged in a line along the posterior margin, and second order ramifications are minute ( Fig. 3A View FIGURE 3 ). Tentacles are in three different sizes, the largest circa 7 mm long, and the smallest usually attached along a line at a more posterior level. The number of tentacles ranges from 26 to 50 in the species; 18 large and medium tentacles and 30 smaller and displaced posteriorly in the holotype. A small distance separates the tentacles and the prepharyngeal groove, and this narrow region is smooth. The prepharyngeal groove is formed by two equal size lamellae that establish a very deep V where the dorsal tubercle is situated. The dorsal tubercle aperture is U or V-shaped, very narrow, lying deep in the V ( Fig. 3B View FIGURE 3 ). The dorsal lamina starts after the insertion of the first right dorsal branchial fold and extends to the esophageal opening with 60–125 long languets, ~ 125 in the holotype. The length of the languets increases posteriorly ( Fig. 3F View FIGURE 3 ).
The pharynx is uncolored and transparent with six high folds per side ( Figs. 2C View FIGURE 2 , 3D View FIGURE 3 ). Fold overlap occurs on both sides, however, the folds on the right side of the holotype show more overlap than the left side. The holotype (11 cm) has 440 total longitudinal vessels and the longitudinal vessel formula is (from right to left):
E 10 (17) 11 (30) 4 (32) 4 (36) 6 (33) 5 (25) 4 DL 3 (35) 10 (30) 7 (33) 8 (32) 9 (35) 10 (16) 5 E
Other samples:
Panama: E 5 (19) 4 (24) 7 (26) 5 (27) 4 (24) 5 (24) 4 DL 2 (21) 4 (22) 7 (26) 6 (24) 6 (23) 6 (24) 5 E
MZUSP 557 (4 cm): E 6 (12) 2 (15) 4 (15) 4 (15) 4 (14) 2 (12) 2 DL 2 (12) 2 (16) 2 (16) 5 (14) 4 (12) 3 (11) 3 E DZUP PYU54 (4.5 cm): E 7 (17) 3 (22) 2 (21) 2 (22) 6 (14) 5 (14) 3 DL 2 (22) 3 (19) 2 (25) 1 (24) 2 (19) 1 (17) 8 E DZUP PYU75 (3.48 cm): E 6 (15) 3(17) 3 (20) 3 (19) 3 (13) 3 (17) 2 DL 2 (16) 1 (15) 2 (19) 2 (18) 3 (17) 2 (12) 4 E MZUSP 0 0 0 41 (3.5 cm): E 8 (10) 4 (16) 4 (16) 3 (16) 3 (16) 3 (15) DL 2 (17) 3 (12) 4 (16) 4 (19) 4 (13) 3 (10) E
There are frequently five or six stigmata per mesh, both between and on folds, but some samples from Brazil had four. Parastigmatic vessels are present; however, not in every mesh and can be incomplete ( Fig. 3E View FIGURE 3 ). Posteriorly languets extend around the right side of the esophageal opening; there are 45 of those languets in the holotype.
The pharynx is attached to the body wall by numerous thick vessels. Significantly smaller ones attach the pharynx to the gonadal and intestinal regions. The intestine is isodiametric, and the primary intestinal loop extends anteriorly, sometimes two thirds to half of the distance to the peripharyngeal groove. The loop is narrow with parallel ascending and descending portions along the ventral margin ( Fig. 2D View FIGURE 2 ). The secondary loop is very small and ends in a smooth anus ( Fig. 3G View FIGURE 3 ). Sometimes, a slight constriction at the pre-anal region could be noted. Endocarps line the ascending and descending intestine, both inside and outside the loop. The ones ventral to the ascending intestine are laminar, very thin and circular, while those lining the descending intestine are fleshy, irregular and more numerous ( Fig. 2D View FIGURE 2 ). Endocarps are also present attached to the body wall. A large digestive gland, 2 cm long in the holotype, shows two connections with the stomach; there are additional small portions on the esophagus ( Fig. 3H View FIGURE 3 ). The atrial velum is red in freshly preserved animals, wide and waving, and without thread-like projections ( Fig. 2D View FIGURE 2 ).
The left gonad occupies the entire space of the first gut loop and contains 34 irregularly shaped lobes in the holotype, but the number of gonad lobes ranges from 34 to 62. They intermittently present fleshy endocarps on the top of the lobes. The right gonad is ventral and takes the form of a backward J. The right gonad extends up to the same position that the intestinal loop does on the left side ( Fig. 2D View FIGURE 2 ), with 62 irregular shaped gonad lobes placed tightly together in the holotype, ranging from 38 to 82 lobes in other samples. The right gonad also has small endocarps on the tops of some lobes. Large and complex fleshy endocarps attached to the body wall line the anterior and dorsal sides of the right gonad. Gonoducts are short on both sides and the oviduct is lobed ( Fig. 3G View FIGURE 3 ).
Remarks. This species can be confused with Pyura vannamei because of the similar color of living animals and the elongated body with a posterior atrial siphon. The distribution of P. vannamei is restricted to the Caribbean region and in the field, it has never been found on mangrove roots. It generally lies horizontally on the ventral side with both siphons up while P. beta generally lies vertically on coral heads or mangrove roots, with the oral siphon up and the atrial siphon posterior and lateral. Pyura beta living color is usually redder and P. vannamei is usually more purple, but both tend to red after preservation in formalin. The two species also differ in that P. beta has colored muscles when fresh and an isodiametric intestine with small secondary loop and smooth anus, whereas P. vannamei has white muscles in fresh samples and an enlarged hind pouch in the second loop, and lobed anus ( Monniot 1994).
Pyura beta View in CoL was first collected from the Brazilian coast in 1961 during an R.V. Calypso expedition and deposited on MZUSP as P. vittata View in CoL by Sérgio A. Rodrigues (Rodrigues 1966). In 1997 one of us (R.M. Rocha) collected two individuals at Cabras Island, São Sebastião, SP, but also identified them as P. vittata View in CoL (DZUP PYU- 22). Since then, many other individuals from the southwestern Atlantic have been collected, mainly from some coastal islands. Brazilian specimens are light reddish to yellow, but all other features are similar with Caribbean ones ( Fig. 4 View FIGURE 4 ). They have been identified as P. vittata View in CoL but both species are different in many aspects: P. vittata View in CoL is more or less globular in shape, the tunic is thin and white internally, the siphons are closer when compared to P. beta View in CoL with long (up to 0.32 mm) and iridescent s pinules noticeable even on the outside of the siphons ( Monniot 2018), and the body musculature is less developed. Also, in P. vittata View in CoL , the right gonad is in a more median portion of the body and it is more dorsally curved ( Monniot 1983).
In a short visit to the Smithsonian Collection in Washington, DC we had the opportunity to see some samples and it is possible that the specimens USNM 21805 and USNM 1090900 collected in Pelican Bay, Belize and identified by I. Goodbody as Pyura lignosa View in CoL may be P. beta View in CoL . External appearance looks exactly the same but there was not enough time for a complete dissection. Goodbody (2000) mentions animals exceeding 15 cm in length. If this identification is confirmed the geographical distribution of the species is enlarged. Monniot (2016) recently reported P. vittata View in CoL from French Guiana but the description given resembles P. beta View in CoL , except for the smaller size and color of preserved animals. The animals do not have the enlarged posterior intestinal region and the author does not mention the presence of siphonal spines, both characteristics typical of P. vittata View in CoL (see Monniot 1983).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pyura beta
| Skinner, Luís Felipe, Rocha, Rosana M. & Counts, Bailey K. 2019 |
Pyura beta
| Skinner & Rocha & Counts 2019 |
P. beta
| Skinner & Rocha & Counts 2019 |
Pyura beta
| Skinner & Rocha & Counts 2019 |
P. beta
| Skinner & Rocha & Counts 2019 |
P. beta
| Skinner & Rocha & Counts 2019 |
Pyura lignosa
| Michaelsen 1908 |