
Pseudorhinolophus Schlosser, 1887
|
publication ID |
https://doi.org/ 10.5252/g2016n3a3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:FC07ACBE-03F7-414A-BB64-1BB0711766BF |
|
persistent identifier |
https://treatment.plazi.org/id/03E07259-FF91-FFA1-FF1D-F975C92DFBC7 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudorhinolophus Schlosser, 1887 |
| status |
|
Sous-genre Pseudorhinolophus Schlosser, 1887
ESPÈCE TYPE. — Pseudorhinolophus morloti Pictet in Pictet et al., 1855 par désignation subséquente ( Schlosser 1887).
AUTRES ESPÈCES DU SOUS- GENRE. — H. (Ps.) dubius Weithofer, 1887; H. (Ps.) weithoferi Revilliod, 1917; H. (Ps.) schlosseri Revilliod, 1917 ; H. (Ps.) egerkingensis Revilliod, 1922; H. (Ps.) bouziguensis Sigé, 1968 ; H. (Ps.) zbrjdi Sigé, 1990; H. (Ps.) russelli Maitre, 2014; H. (Ps.) major Maitre, 2014; H. (Ps.) tenuis Maitre, 2014.
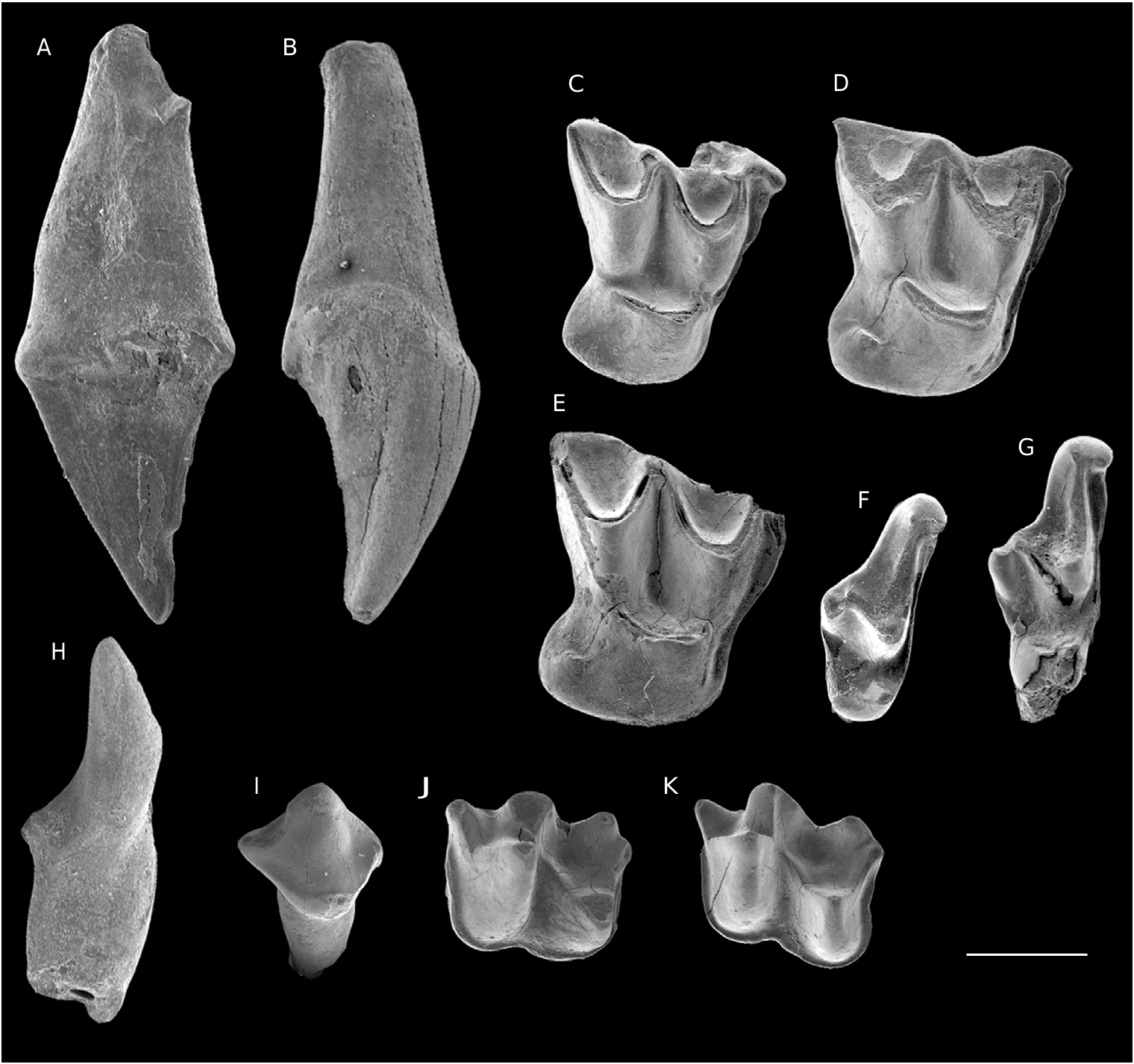
Hipposideros (Pseudorhinolophus) africanum Ravel , n. sp. ( Figs 8 View FIG , 9 View FIG , 10 View FIG ; Tableau 3)
HOLOTYPE. — Spécimen CBI-1-247, fragment de maxillaire droit avec M2 et M3; Figs 8C View FIG ; 9 C. View FIG
MATÉRIEL EXAMINÉ. — CBI-2-045 ( Figs 8A View FIG ; 9 A View FIG ), CBI-2-047 ( Fig. 10B View FIG ) et CBI-2-048 (C1s droites; Fig. 10A View FIG ); CBI-2-046 ( Figs 8B View FIG ; 9 B View FIG ) et CBI-2-049 (C1s gauches); CBI-1-248 (M1 droite; Figs 8D View FIG ; 9 D View FIG ); CBI-2-039 ( Fig. 10C View FIG ), CBI-2-043 ( Figs 8E View FIG ; 10D View FIG ) et CBI-2-044 (M2s droites; Fig. 10E View FIG ); CBI-2-041 ( Fig. 10G View FIG ) et CBI-2-042 (M3s droites; Fig.10F View FIG ); CBI-2-040 (M3 gauche); CBI- 2-053 (fragment de dentaire droite avec m2 et m3; Figs 8F View FIG ; 9 G View FIG ); CBI-2-057 ( Figs 8H View FIG ; 9 E View FIG ) et CBI-2-058 (c1s droites; Fig. 10H View FIG ); CBI-2-056 (c1 gauche; Figs 8I View FIG ; 9 F View FIG ); CBI-2-054 ( Figs 8G View FIG ; 9 H View FIG ) et CBI-2-055 (p2s droites; Fig. 10I View FIG ); CBI-2-220 et CBI-3-004 (p2 gauche; Fig.10 I View FIG ); CBI-2-050 ( Figs 8J View FIG ; 10 K View FIG ) et CBI-2-051 (m1/2s droites; Fig. 9 K View FIG ); CBI-1-249 ( Fig. 10J View FIG ), CBI-2-250 ( Figs 8K View FIG ; 9 J View FIG ), et CBI-2-052 (m1/2s gauches; Fig. 9L View FIG ).
ÉTYMOLOGIE. — Le nom de l’espèce fait référence au continent africain et souligne l’originalité géographique de l’espèce qui appartient à un genre qui était documenté exclusivement en Europe.
LOCALITÉ TYPE ET ÂGE. — Chambi (CBI) loci 1, 2 et 3, Éocène inférieur terminal-Éocène moyen basal, Djebel Chambi, situé dans la région de Kasserine, Tunisie.
DIAGNOSE. — Diffère des autres espèces d’ Hipposideros (Pseudorhinolophus) par le faible développement et l’inclinaison distale du mésostyle, l’inclinaison mésiolinguale du bord buccal de M1, le cingulum lingual très épais et peu étendu, le développement à prédominance transversale de M3, et l’important développement longitudinal du talonide.
A C
E D
DESCRIPTION
Les canines supérieures ont une morphologie simple ( Figs8A, B View FIG ; 9A, B View FIG ; 10A, B View FIG ). L’unique tubercule est peu élevé, robuste, et montre un apex incliné distalement. La face buccale de la cuspide principale est bombée et s’oppose à une face linguale plane. Le cingulum est bien individualisé lingualement mais est absent sur la face buccale. Il forme à la base distale de la couronne un petit méplat (CBI-2-045 et CBI-2-046), ou forme un ressaut cuspidé relié par la crête distale du tubercule (CBI- 2-047 et CBI-2-049). La canine CBI-2-048 présente un cône moins incliné et une taille plus importante. Cette différence pourrait être liée soit à une variation intraspécifique, soit à la présence d’une autre espèce proche. Le manque de données ne permet pas de pencher pour l’une ou l’autre des hypothèses. La première et la seconde molaire supérieure présentent de nettes différences morphologiques. La M1 est très compressée buccolingualement, ce qui lui confère un contour occlusal plutôt quadrangulaire ( Figs 8D View FIG ; 9 D View FIG ). Son bord buccal est incliné mésiolingualement et effectue un léger ectoflexus au dessus du paracône. Le parastyle est courbé vers l’avant, et apparait dès lors en position plus linguale que le métastyle, ce dernier étant plutôt rectiligne. Le « V» antérieur de l’ectolophe, formé par la pré- et post- paracrêtes, est plus ouvert que le «V» postérieur (formé par les pré- et post- métacrêtes). Le mésostyle est peu développé et s’incline distalement sans se projeter au-delà du bord buccal. Le paracône est moins volumineux et plus lingual que le métacône. Le protocône est très effilé (comprimé buccolingualement) par rapport aux deux tubercules buccaux. Il est légèrement plus distal que le paracône. La préprotocrête s’étend buccalement jusqu’à la base du paracône, délimitant ainsi la paroi mésiale de la couronne. Le précingulum, très court, s’initie buccalement depuis la base du parastyle et s’étend distalement jusqu’à l’aplomb du paracône. La postprotocrête est environ deux fois plus longue que la préprotocrête. La postprotocrête s’étend vers la partie distale de manière quasi horizontale en vue occlusale. Elle s’arrête à la base linguale du métacône, laissant ainsi la protofosse ouverte distalement. Un cingulum lingual apparaît dans la région distolinguale de la couronne et prend la forme d’un épais lobe constituant le talon de la molaire.
Comparées à la M1, les M2s sont plus développées transversalement, avec un contour occlusal plus rectangulaire ( Figs 8 View FIG C-E; 9C, D; 10C-E). Le bord buccal est presque horizontal selon le plan occlusal, avec un léger ectoflexus au dessus du paracône (plus prononcé sur le spécimen CBI-2- 039). L’ectolophe de M2 est plus symétrique et plus large que celui de M1. Le parastyle est plus déjeté buccalement, se plaçant ainsi sur le même axe mésiodistal que le métastyle. Le mésostyle est réduit et légèrement en retrait du bord buccal. Le protocône est très compressé buccolingualement et moins allongé mésiodistalement que sur M1. Une partie du cingulum lingual est visible à la base du flanc mésial du protocône. Le lobe distolingual de M2 est moins projeté vers l’arrière que sur M1. Un postcingulum vestigial s’étend depuis la base du métastyle jusqu’au point d’inflexion du lobe distolingual.
Les M3s ont une largeur équivalente aux M2s ( Figs 8C View FIG ; 9C View FIG ; 10 F, G View FIG ). Ces molaires sont toutefois très courtes. Le bord buccal est très incliné distolingualement et infléchit au niveau du métacône. Le métacône est très peu développé et constitue un petit renflement distal. La prémétacrête est très courte tandis que la postmétacrête est absente. La partie linguale est dominée par le protocône dont le volume est inférieur à celui du paracône. Les protocrêtes sont très courtes et isolées. Le bord lingual arrondi est dépourvu de cingulum.
Deux molaires supérieures présentent une variation très forte. CBI-2-043 ( Figs8E View FIG ; 10D View FIG ) et CBI-2-044 ( Fig.10E View FIG ) possèdent un très large précingulum s’étendant sur toute la largeur mésiale de la couronne depuis le parastyle. Cette structure est peu développée chez les autres spécimens. Cependant cette variation pourrait être due à une variation intraspécifique, mais ne peut, à elle seule, justifier la présence d’une autre espèce. Les canines inférieures se composent d’un unique tubercule très légèrement recourbé ( Figs 8H, I View FIG ; 9E, F View FIG ; 10 H View FIG ). Le cingulum est bien marqué sur la face linguale tout en étant incliné distalement. Il forme un léger relief à la base mésiale de la cuspide et cerne un long bassin qui s’étend à la base distale du tubercule. Le cingulum est absent au niveau du bord buccal. La deuxième prémolaire possède une couronne à contour ovale ( Figs 8G View FIG ; 9H, I View FIG ; 10I View FIG ). Elle se caractérise par le développement d’une seule cuspide, lequel constitue la majeure partie de la dent. Ce dernier est peu élevé, à base très large, et possède deux crêtes sagittales séparant la face buccale de la face linguale. La face buccale est légèrement plus bombée que la face linguale. Le cingulum, épais et continu, s’incurve buccalement. Un petit relief apparaît à la bordure distolinguale de la couronne formé en partie par un ressaut du cingulum.
Le talonide des molaires inférieures est très légèrement plus long que le trigonide ( Figs 8F, J, K View FIG ; 9G, J, K, L View FIG ; 10J, K View FIG ). Le trigonide est plutôt comprimé mésiodistalement: la distance entre le paraconide et le métaconide est moins importante que celle entre le protoconide et le paraconide ou le métaconide. Le paraconide est la cuspide du trigonide la plus réduite, il est également proclive. Le protoconide est droit, son flanc buccal est pincé mésiodistalement. Le talonide est plus large que le trigonide mais moins élevé en hauteur. L’hypoconide est le tubercule dominant du talonide. Il se situe plus buccalement que le protoconide. L’entoconide, de hauteur modérée par rapport aux autres tubercules du talonide, est légèrement plus distal que l’hypoconide. L’entocristide forme une encoche linguale entre l’entoconide et le métaconide. L’hypoconulide est proche distobuccalement de l’entoconide. Un petit hypoconulide, plus réduit que l’entoconide, est directement connecté à l’hypoconide par la postcristide dessinant ainsi une structure nettement nyctalodonte. La crête oblique commence depuis la base distale du trigonide entre le protoconide et le métaconide et rejoint la pointe de l’hypoconide. Le cingulide, modérément marqué, est continu sur tous les bords mésial, buccal et distal de la dent.
La m3 est plus petite que les m1s et m2s ( Figs 8F View FIG ; 9G View FIG ). Le trigonide de m3 n’est pas très modifié et se différencie par une taille plus petite et un resserrement mésiodistal du métaconide et du paraconide. La réduction de m3 est surtout réalisée au niveau du talonide. Cependant les trois cuspides du talonide sont bien visibles et de tailles similaires. L’hypoconulide est très projeté distalement à l’inverse de l’entoconide, lequel forme un petit bourrelet près de la base distale du métaconide. Le cingulide est également continu mais rétréci fortement au niveau du bord mésial et se termine buccalement à l’hypoconide.
COMPARAISON ET DISCUSSION
Ces fossiles de Chambi possèdent un ensemble de caractères qui se rencontrent plus généralement chez les Hipposideridae , et plus précisément du genre Hipposideros . Il s’agit notamment de:
– l’aspect comprimé buccolingualement des molaires supérieures;
– la morphologie de l’ectolophe;
– l’exiguïté de la protofosse;
– la proportion du protocône;
– l’extension du talon (lobe);
– des canines supérieures courbées et dépourvues de cingulum buccal;
– la présence d’un entoconide faible;
– la réduction latérale de la M3 (plus courant chez les Hipposideridae que chez les Rhinolophidae );
– la réduction du talonide de la m3 (également plus courant chez les Hipposideridae que chez les Rhinolophidae );
– l’extension de la postprotocrête.
Le genre Hipposideros est abondamment documenté dans les poches karstiques du Quercy depuis l’Éocène moyen terminal (Saint Lizier, MP16; une espèce reste non identifiée au niveau MP13 à Chamblon), jusqu’au début de l’Oligocène supérieur (Garouillas, MP25) et au Néogène ( Sigé 1978, 1990; Sigé & Legendre 1983; Maitre 2014). Ces fossiles ont été regroupés dans un sous-ensemble Hipposideros (Pseudorhinolophus) , parfois utilisé comme genre à part entière ( Hand 1998a, b) et dont la nomination rappelle les affinités morphologiques avec le genre Rhinolophus . Hipposideros (Pseudorhinolophus) africanum Ravel , n. sp. de Chambi partage avec les formes européennes:
– l’absence de cingulum buccal sur les canines;
– des faces lisses au tubercule des canines supérieures;
– le développement des crêtes internes de l’ectolophe;
– un paracône et un métacône de taille équivalente;
– la projection distale du talon de M1 plus importante par rapport à celle sur M2;
– la réduction de M3, avec la présence d’une arête distale au mésostyle;
– la présence d’un relief distolingual sur p2 (rudiment de paraconide);
– la réduction du talonide de m3.
L’ensemble de ces traits contribue à caractériser la morphologie dentaire des Hipposideros (Pseudorhinolophus) , confirmant ainsi l’appartenance du taxon tunisien à ce sous-groupe. Hipposideros (Ps.) africanum Ravel, n. sp. possède également certaines divergences morphologiques avec les formes européennes, soulignant son identité spécifique:
– mésostyle moins projeté buccalement et incliné distalement;
– forte inclinaison mésiolinguale du bord buccal de M1;
– M3 relativement plus développée transversalement;
– épaisseur du cingulum lingual sur M1-2 plus importante avec une extension mésiale plus réduite;
– extension du précingulum le long de la bordure mésiale de la couronne (variable parmi le matériel référé à H. [Ps.] africanum Ravel , n. sp.);
– tubercules des molaires inférieures mieux individualisés.
Hipposideros (Pseudorhinolophus) africanum Ravel , n. sp. témoigne de la présence de l’ensemble Pseudorhinolophus dès la fin de l’Éocène inférieur en Afrique. Les espèces européennes du groupe ne sont documentées qu’à partir de la fin de l’Éocène moyen. Cette espèce représente donc l’occurrence la plus précoce. Il est possible que le taxon tunisien, de par son ancienneté et son originalité vis-à-vis des autres espèces du même genre et sous-genre, reflète une évolution de la morphologie dont les tendances seraient au sein de cet ensemble: développement du mésostyle, redressement du bord buccal de M1, affinement et extension du cingulum lingual, réduction de M3, renforcement des cuspides des molaires inférieures, raccourcissement du talonide. Cependant le matériel reste insuffisant pour préciser les modalités macroévolutives des formes Hipposideros .
K
L M N
Super-famille EMBALLONUROIDEA Dobson, 1875 Famille EMBALLONURIDAE Gervais in de Castelnau, 1855


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |