
Paraleochara kochi Bernhauer, 1941
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4101.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:53A25A6F-72AF-4B61-B8DA-E427FA2B634F |
|
DOI |
https://doi.org/10.5281/zenodo.6056052 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87F7-FFE7-0D5F-FF21-FFA4FB2DFEBC |
|
treatment provided by |
Plazi |
|
scientific name |
Paraleochara kochi Bernhauer, 1941 |
| status |
comb. nov. |
Paraleochara kochi Bernhauer, 1941 View in CoL , n. comb.
( Figs 9 View FIGURES 1 – 9 , 18 View FIGURES 10 – 18 , 27 View FIGURES 19 – 27 , 36 View FIGURES 28 – 36 , 44 View FIGURES 37 – 44 , 100–106 View FIGURES 100 – 106 , 119 View FIGURES 107 – 119 , 128 View FIGURES 127 – 128 )
Aleochara (Polychara) kochi Bernhauer, 1941: 211 View in CoL [original description; type locality: Japan, “Umgebung Tokio” (around Tokyô-to, Honshû)].
Aleochara (Aleochara) kochi Bernhauer, 1941 View in CoL ; Likovský, 1973: 2 (taxonomic placement; notes on Palearctic Aleochara View in CoL ); Smetana, 2004b: 354 (catalogue of Palearctic species of Aleocharinae View in CoL ); Park & Ahn, 2010a: 313 (redescription), 317 (in key to Far-Eastern species of subgenus Aleochara View in CoL ); Shibata et al., 2013: 105 (catalogue of Japanese species of Staphylinidae View in CoL ); Schülke & Smetana, 2015: 491 (catalogue of Palearctic species of Aleocharinae View in CoL ).
Aleochara globus Pace, 1999: 158 (as Aleochara View in CoL (s. str.); original description; type locality: China: Zhejiang, Lin’an County); Smetana, 2004b: 354 (as subgenus Aleochara View in CoL ; catalogue of Palearctic species of Aleocharinae View in CoL ); Luo & Zhao, 2012: 180 [as Aleochara View in CoL (s. str.); in key to Chinese species of subgenus Aleochara View in CoL ], 195 (redescription); Schülke & Smetana, 2015: 491 (as subgenus Aleochara View in CoL ; catalogue of Palearctic species of Aleocharinae View in CoL ). New synonymy
See mouthparts description for Park & Ahn (2010a).
Type material. A. kochi : Lectotype, male, “ Japonia: Umg. / Tokyo, VI.–VIII. 25 // Aleochara / kochi Brnh / n. sp. [HW] // kochi Bernh / Typus / Polychara [RPC, HW] // Chicago NHMus / M. Bernhauer / Collection // LECTOTYPE / Aleochara kochi / Bernhauer, 1941 / Desig. Park & Ahn, 2007 [RPC]” (aedeagus, parameres and mouth parts dissected and mounted by PR) (BL, ≈ 4.9 mm; PL, 0.80 mm; PW, 1.08 mm; HTL, 0.90 mm) ( FMNH).
A. globus: Not examined.
Non-type material. JAPAN: [HOKKAIDÔ]: 1 unsexed, Nopporo, Ebetsu-shi, 17.vi.1986, M. Ôhara leg., from baited trap ( HUM); 1 unsexed, Nagahashinaebo, Otaru-shi, 31.v.1991, M. Ôhara leg. ( OCM); 3 unsexed, Nagahashinaebo, Otaru-shi, 15.v.1992, M. Ôhara leg., from baited trap ( OCM); 3 unsexed, Nagahashinaebo, Otaru-shi, 9.v.1992, M. Ôhara leg., from baited trap ( OCM); 1 unsexed, Nagahashinaebo, Otaru-shi, 25.vi.1992, M. Ôhara leg., from baited trap ( OCM); 1 unsexed, Junsai-numa, Nanae-chô, 7.vii.1918, M. Suzuki leg. ( ELKU); [HONSHÛ]: 1 unsexed, Yoriguri ~ Kamikuga, Kanuma-shi, Tochigi-ken, 21.vii.1997, H. Ôkawa leg. (KUM); 1 male, 2 unsexed, Mt. Arima-yama, Hannô-shi, Saitama-ken, 17–24.vii.2004, K. & S. Arai leg., alt. 1200m (KUM); 1 unsexed, Motoishikawa, Aoba-ku, Yokohama-shi, Kanagawa-ken, 2.iii.2002, H. Ooki leg. (cHO); 1 unsexed, Yokohama National University, Tokiwadai, Hodogaya-ku, Yokohama-shi, Kanagawa-ken, 19.iv.2010, H. Ooki leg. (cHO); 2 unsexed, Ôwakudani, Hakone-machi, Kanagawa-ken, 7–17.viii.2007, T. Watanabe leg., by flight interception trap (cTW); 2 unsexed, Ôwakudani, Hakone-machi, Kanagawa-ken, 17–27.viii.2007, T. Watanabe leg., by flight interception trap (cTW); 1 male, Nakatsu, Aikawa-machi, Kanagawa-ken, 2–12.iii.2010, T. Watanabe leg., from chicken baited trap near river (cTW); 4 unsexed, Nakatsu, Aikawa-machi, Kanagawa-ken, 22.ii–12.iii.2010, T. Watanabe leg., from chicken baited trap near river (cTW); 1 male, 3 unsexed, Karumizu-rindô forestry road, Narusawa-mura, Yamanashi-ken, 1–8.viii.2012, T. Watanabe leg., alt. 1600m, from flight interception trap (cTW); 1 male, 2 unsexed, Karumizu-rindô forestry road, Narusawa-mura, Yamanashi-ken, 3– 10.viii.2012, T. Watanabe leg., alt. 1600m, from flight interception trap (cTW); 1 male, 4 unsexed, Karumizu-rindô forestry road, Narusawa-mura, Yamanashi-ken, 10–17.viii.2012, T. Watanabe leg., alt. 1600m, from flight interception trap (cTW); 1 male, 3 unsexed, Mt. Takahachi-yama, Fujinomiya-shi, Shizuoka-ken, 16–22.vii.2010, T. Watanabe leg., from flight interception trap (cTW); 2 males, Mt. Takahachi-yama, Fujinomiya-shi, Shizuokaken, 31.vii–6.viii.2010, T. Watanabe leg., from flight interception trap (cTW); 2 males, 1 female, Mt. Takahachiyama, Fujinomiya-shi, Shizuoka-ken, 17–24.viii.2010, T. Watanabe leg., from flight interception trap (cTW); 1 unsexed, Mt. Takahachi-yama, Fujinomiya-shi, Shizuoka-ken, 24.viii.2010, T. Watanabe leg. (cTW); 2 males, 3 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 16–22.vii.2010, T. Watanabe leg., from flight interception trap (cTW); 2 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 31,vii– 6.viii.2010, T. Watanabe leg., from flight interception trap (cTW); 5 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 17–24.viii.2010, T. Watanabe leg., from flight interception trap (cTW); 4 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 22– 28.viii.2010, T. Watanabe leg., from baited trap (silkworm pupa powder) (cTW); 2 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 28.viii–5.ix.2010, T. Watanabe leg., from baited trap (silkworm pupa powder) (cTW); 1 unsexed, Nishiusuzuka, Fujinomiya-shi, Shizuoka-ken, 14–21.ix.2010, T. Watanabe leg., from flight interception trap (cTW); 1 unsexed, Ôbuchi, Fuji-shi, Shizuoka-ken, 20.iv.1999, Y. Hirano leg. (KUM); 1 female, Mt. Amaishi-yama, Sasayama-shi, Hyôgo-ken, 2.v.1980, Y. Hayashi leg. (cYH); 3 unsexed, Sasayama-shi, Hyôgoken, 26.iv.1980, Y. Hayashi leg. (cYH); 1 unsexed, Takiyama, Nagi-chô, Okayama-ken, 27.iv.2003, Y. Fujitani leg., alt. 500–700m, from pitfall trap (KUM); 5 unsexed, Tokujinotani, Yamaguchi-shi, Yamaguchi-ken, 10.iv.2004, Y. Fujitani leg. (KUM); 1 unsexed, Tokujinotani, Yamaguchi-shi, Yamaguchi-ken, 15.iv.2004, Y. Fujitani leg. (KUM); 5 females, 15 unsexed, Tokujinotani, Yamaguchi-shi, Yamaguchi-ken, 21.iv.2004, Y. Fujitani leg. (KUM); [SHIKOKU]: 1 unsexed, Mt. Rokuro-yama, Sanagôchi-son, Tokushima-ken, 5.v–1.vii.2000, M. Yoshida leg., alt. 830m, from fowl trap (KUM); 1 unsexed, Mt. Maruzasa-yama, Tsurugi-chô, Tokushima-ken, 11.vi–6.viii.2000, M. Yoshida leg., alt. 1410m, from fowl trap (KUM); 1 unsexed, Nishijima (Mt. Tsurugi-san), Higashiiya, Miyoshi-shi, Tokushima-ken, 26–29.vii.2006, M. Yoshida leg., alt. 1700m, from fowl trap (KUM); 1 unsexed, Ôtsurugi (Mt. Tsurugi-san), Higashiiya, Miyoshi-shi, Tokushima-ken, 5.vi.–3.vii.2006, M. Yoshida leg., alt. 1800m, from fowl trap (KUM).
Diagnosis. Paraleochara kochi can be separated from P. fungivora by having strongly narrowing ligula and by much developed apical invagination of female spermatheca ( Sawada, 1982; Park & Ahn, 2010a). This species also externally resembles Tetrasticta laeta Maruyama & Sugaya, 2002 from the Ryûkyûs ( Japan) and Taiwan, but it can be distinguished from T. laeta as follows: body much slender, parallel sided; pronotum without prominent three pairs of characteristically arranged punctation; elytra with blackish maculations laterally, but much smaller and indistinct; abdominal tergite VIII of male with serrate posterior margin; abdominal sternite VIII sexually dimorphic, with sharply pointed apex in male; median lobe of male aedeagus without coiled flagellum; female spermatheca different in shape, with huge spherical apical invagination of spermathecal head. See descriptions and illustrations of T. laeta in Maruyama & Sugaya (2002). From all of the Japanese species of the genus Aleochara , P. kochi can be easily distinguished by having very short, widely truncated, intercoxal process of mesoventrite ( Fig. 36 View FIGURES 28 – 36 ).
Redescription. Measurements (in mm, n = 20): BL = 4.99 (3.96–5.88); HL = 0.66 (0.58–0.75); HW = 0.76 (0.67–0.83); PL = 0.84 (0.70–0.95); PW = 1.12 (0.96–1.22); EL = 0.85 (0.70–1.00); EW = 1.31 (0.90–1.52).
Body ( Figs 9 View FIGURES 1 – 9 , 18 View FIGURES 10 – 18 ): rather subparallel sided, slender, small to medium sized; dorsal surface strongly glossy and roughly pubescent, without any types of micro-reticulation, finely and shallowly punctured. Color ( Figs 9 View FIGURES 1 – 9 , 18 View FIGURES 10 – 18 ): usually uniformly dark reddish brown to brownish brown, but elytra bright yellowish brown, ambiguously infuscate posterolaterally, not reaching to shoulders of each elytron; antennae with antennomeres I–III dark yellowish brown, antennomere IV darker, antennomeres V–XI much darker than those of I–III but densely covered with minute whitish setae; mouth parts and legs pale yellowish brown to reddish brown; pubescence yellowish brown.
Head ( Fig. 27 View FIGURES 19 – 27 ): circular, as long as width (HW/HL = 1.14, n = 20), widest at posterior part of eyes; setae on vertex rather sparse but long, directed anteriomedially; punctation medium in size but shallow, scattered sparsely; vertex longitudinally lack of punctation along midline, but small to medium in size. Eyes: relatively long, occupying more than half length of head, very slightly protruding laterally, almost incorporated to head capsule.
Antennae ( Fig. 44 View FIGURES 37 – 44 ): medium in length, slightly shorter than head and pronotum combined; relatively thick, setose; antennomere IV strongly transverse, antennomeres V to X strongly transverse in approximately same width each, antennomere XI semicircular, short, clearly shorter than antennomeres IX and X combined; approximate relative length of antennomeres from basal to apex: 19 (including base): 10: 10: 3: 5: 5: 5: 5: 5: 6: 12.
Pronotum ( Fig. 9 View FIGURES 1 – 9 ): transversely oval (PW/PL = 1.33, n = 20), slightly shorter than sutural length of elytra, widest at middle, basal margin moderately rounded; pubescence rather dense, directed laterally and posterolaterally, with approximately seven blackish bristles situated along with each lateral margin; punctuation small and shallow, covered with sparsely and uniformly.
Mesoventrite ( Fig. 36 View FIGURES 28 – 36 ): inter coxal process very short, with widely truncate apex, completely reaching to inter coxal process of metaventrite.
Metaventrite ( Fig. 36 View FIGURES 28 – 36 ): inter coxal process of metaventrite extremely long, narrowly elongate above, with rather widely truncate apex.
Elytra ( Fig. 9 View FIGURES 1 – 9 ): together, transverse (EW/EL = 1.54, n = 20), moderate in size, slightly larger than pronotum, widest around posterior half; dorsal surface with fine, minute, inconspicuous pubescence scattered densely, diverging posteriorly in each elytron; dorsal surface very slightly rough and weakly impressed; posterolateral corners of each elytron not sinuate.
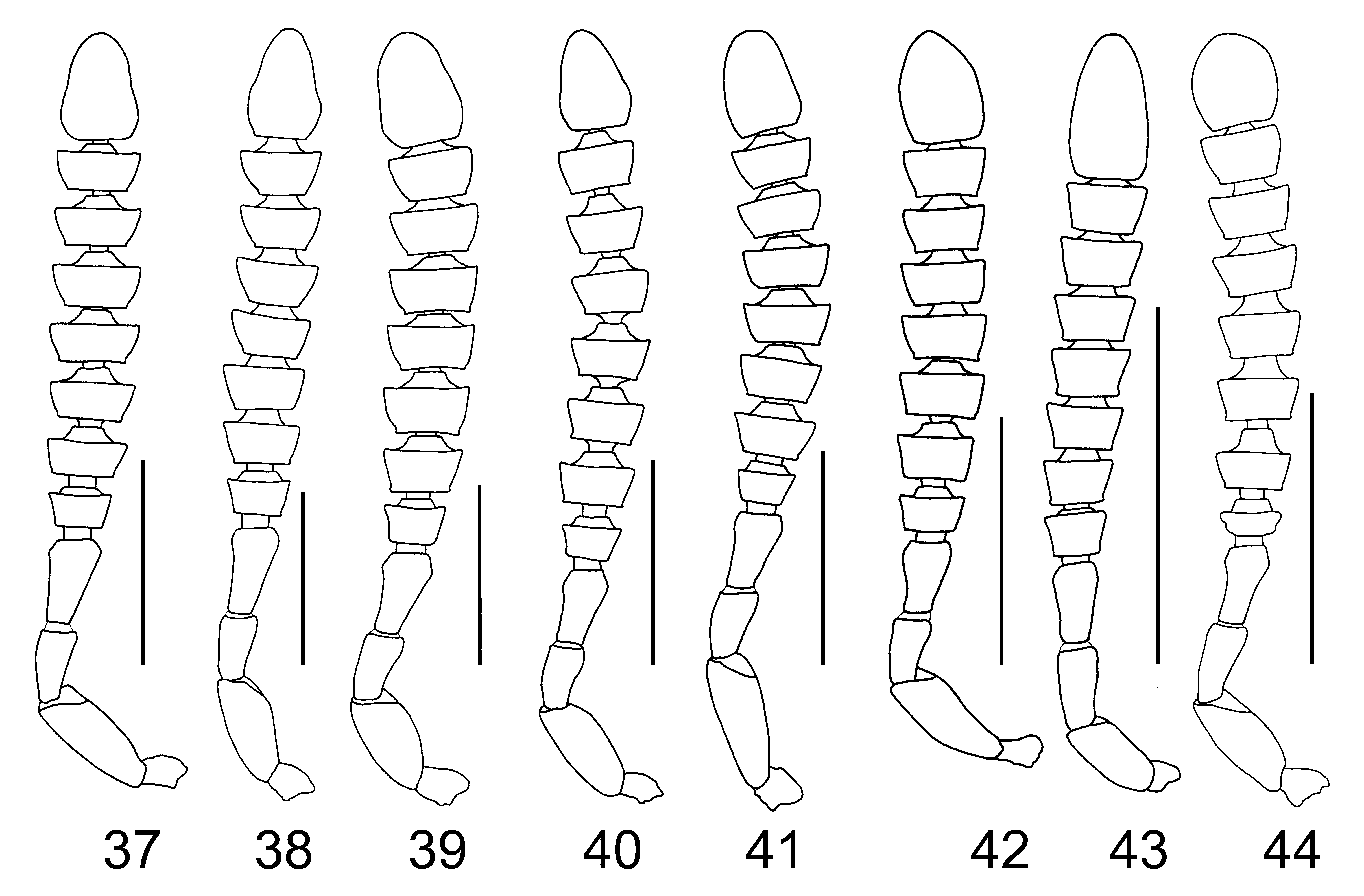
Legs ( Fig. 9 View FIGURES 1 – 9 ): simple, long and slender; fore and midtibia, densely covered with undeveloped small spines, respectively.
Abdomen ( Fig. 9 View FIGURES 1 – 9 ): first three visible tergites deeply impressed transversely at base; dorsal and ventral surface coarsely covered with long, thin setae.
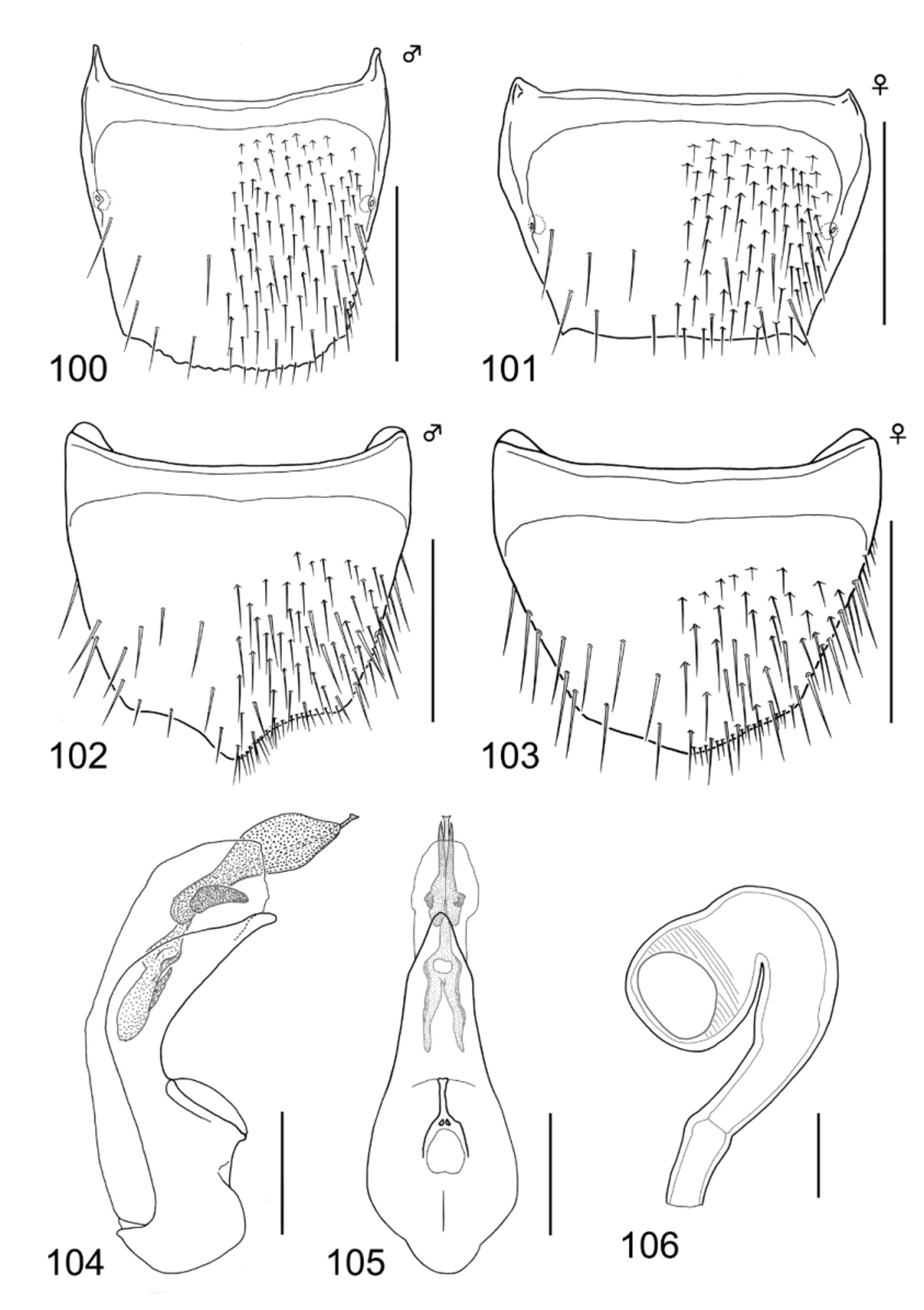
Male: tergite VIII ( Fig. 100 View FIGURES 100 – 106 ): posterior margin moderately but clearly serrate; dorsal surface covered with setae moderately, with approximately six macrosetae. Sternite VIII ( Fig. 102 View FIGURES 100 – 106 ): posterior margin sharply pointed, with a dense row of minute but relatively long sensory setae (see Yamamoto & Maruyama, 2012: 10); no complex-cluster of minute setae situated near posterior margin; ventral surface coarsely covered with setae, with approximately 12 macrosetae. Median lobe of aedeagus ( Figs 104–105 View FIGURES 100 – 106 ): [lateral view ( Fig. 104 View FIGURES 100 – 106 )]: robust, moderately narrowing apically; apical lobe pointed, not hook-like shaped at apex; basal swelling of median lobe small, not strongly developed; [parameral view ( Fig. 105 View FIGURES 100 – 106 )]: elongate triangular, with basal part compactly rounded; apical lobe abruptly rather weakly narrowing near apex, forming obtusely pointed apex; sclerites simple, restricted to apical part, with a projecting short flagellum, as half as median lobe.
Female: tergite VIII ( Fig. 101 View FIGURES 100 – 106 ): posterior margin smooth, not serrate at all; dorsal surface covered with setae moderately, with approximately six macrosetae. Sternite VIII ( Fig. 103 View FIGURES 100 – 106 ): posterior margin moderately pointed; ventral surface covered with setae coarsely, with approximately 11 long macrosetae; no complex-cluster of minute setae situated near posterior margin. Spermatheca ( Fig. 106 View FIGURES 100 – 106 ): deformed and folded C-shaped; apical invagination of spermatheca spherical and extremely large, located deeply inward; spermathecal head and neck weakly semicircular together; attachment of spermathecal duct inconspicuous; basal part of spermathecal stem gently curved, longer than spermathecal head and neck combined; chitinized portion of spermathecal stem relatively short in length, shorter than spermathecal head and neck combined; each part of spermatheca uniformly and weakly sclerotized; inner wall of spermathecal head coarsely striate, that of basal part not reticulate or striate at all.
Distribution. Japan (Hokkaidô, Honshû, Shikoku); Korea, China.
Bionomics. No detailed bionomics have been reported to date. Several specimens were caught by flight interception traps.
Remarks for type material. A single specimen of “ A.” kochi ( Fig. 119 View FIGURES 107 – 119 ) is preserved in FMNH as the lectotype, a designation made by Park & Ahn (2010a).
Comments. In this paper, we transfer “ Aleochara ” kochi to the genus Paraleochara , in the Oriental region, based on its generic morphological characteristics, e.g., prominently glossy body, very short intercoxal process of mesoventrite with a broadly truncated posterior margin, and long and slender legs ( Cameron, 1920, 1939; Sawada, 1982). Paraleochara kochi n. comb. is a second species of the genus. Furthermore, the occurrence of the genus in Japan represents a new distribution record, even for the Palearctic region.
Pace (1999) described “ Aleochara (s. str.)” globus , based on one female specimen from the mainland of China.
Subsequent authors followed the taxonomic treatment, maintaining the species validity ( Smetana, 2004b; Luo & Zhao, 2012; Schülke & Smetana, 2015). Diagnostic characteristics including the structure of female spermatheca ( Pace, 1999), however, were not sufficient to maintain the separation of “ A.” globus and P. kochi . Here, we therefore synonymize “ A.” globus n. syn. under P. kochi .
The distribution of P. kochi is wide in Japan but is restricted only to the mainland ( Fig. 128 View FIGURES 127 – 128 ).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paraleochara kochi Bernhauer, 1941
| Yamamoto, Shûhei & Maruyama, Munetoshi 2016 |
Aleochara globus
| Schulke 2015: 491 |
| Luo 2012: 180 |
| Smetana 2004: 354 |
| Pace 1999: 158 |
Aleochara (Aleochara) kochi
| Schulke 2015: 491 |
| Shibata 2013: 105 |
| Park 2010: 313 |
| Smetana 2004: 354 |
| Likovsky 1973: 2 |
Aleochara (Polychara) kochi
| Bernhauer 1941: 211 |