
Paepalanthus leucocephalus Ruhland, 1903
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.560.2.1 |
|
DOI |
https://doi.org/10.5281/zenodo.7046982 |
|
persistent identifier |
https://treatment.plazi.org/id/038687A1-FFF7-9547-9FE8-225DFB997EA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Paepalanthus leucocephalus Ruhland |
| status |
|
3. Paepalanthus leucocephalus Ruhland View in CoL in Engler, Pflanzenr. IV.30 (Heft 13): 200. 1903.
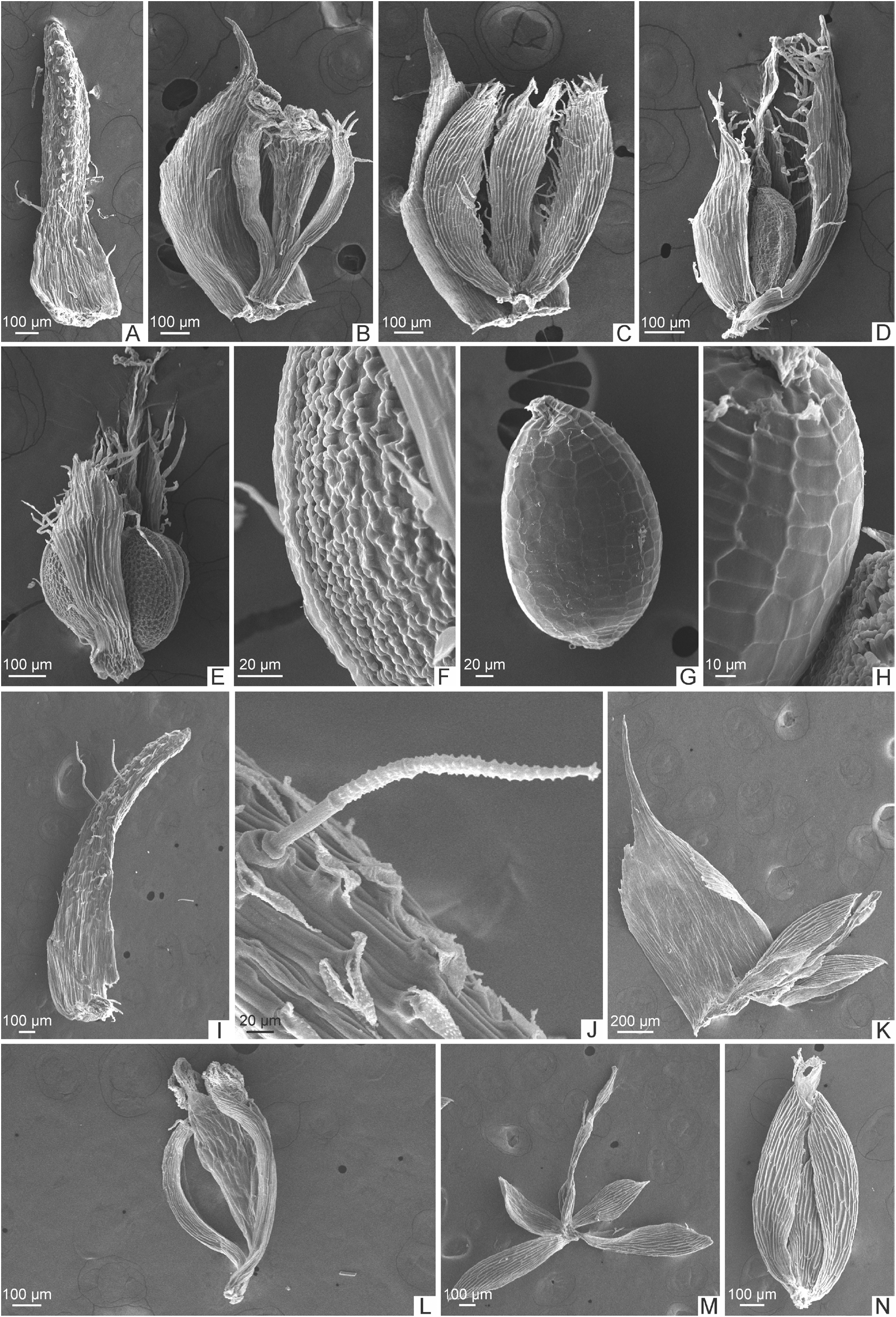
Lectotype, first-step (designated by Moldenke 1975): at B. Lectotype, second-step (here designated): BRAZIL. Minas Gerais: Serra dos Cristaes , 4 April 1892, C.A.W. Schwacke 8503 ( B [10_0243944]! , isolectotypes B [10_0243943]!, K [000640071]!). ( Figures 2A–B View FIGURE 2 , 3 View FIGURE 3 , 4N View FIGURE4 –P, 6A–H View FIGURE 6 )
Ruhland (1903) described Paepalanthus leucocephalus based on the collection Schwacke 8503, gathered in Minas Gerais. We found three specimens related to this collection, two deposited at B, where Willy Ruhland mostly worked, and one housed at K. It is also a situation with no holotype being clearly indicated, requiring a lectotype selection ( McNeill 2014, Turland et al. 2018). Moldenke (1975) indicated that the type of this species is deposited at B with the indication “ Thelxinoë leucocephala ”. The two specimens at B contain the same annotation, therefore we consider Moldenke’s (1975) statement as a first-step lectotype indication according to the article 9.17 of the ICN ( Turland et al. 2018). One of the specimens housed at B contains Ruhland’s original handwritings and drawings attached, also perfectly matching the description. We designate this specimen [B barcode 10_0243944] as the second-step lectotype.
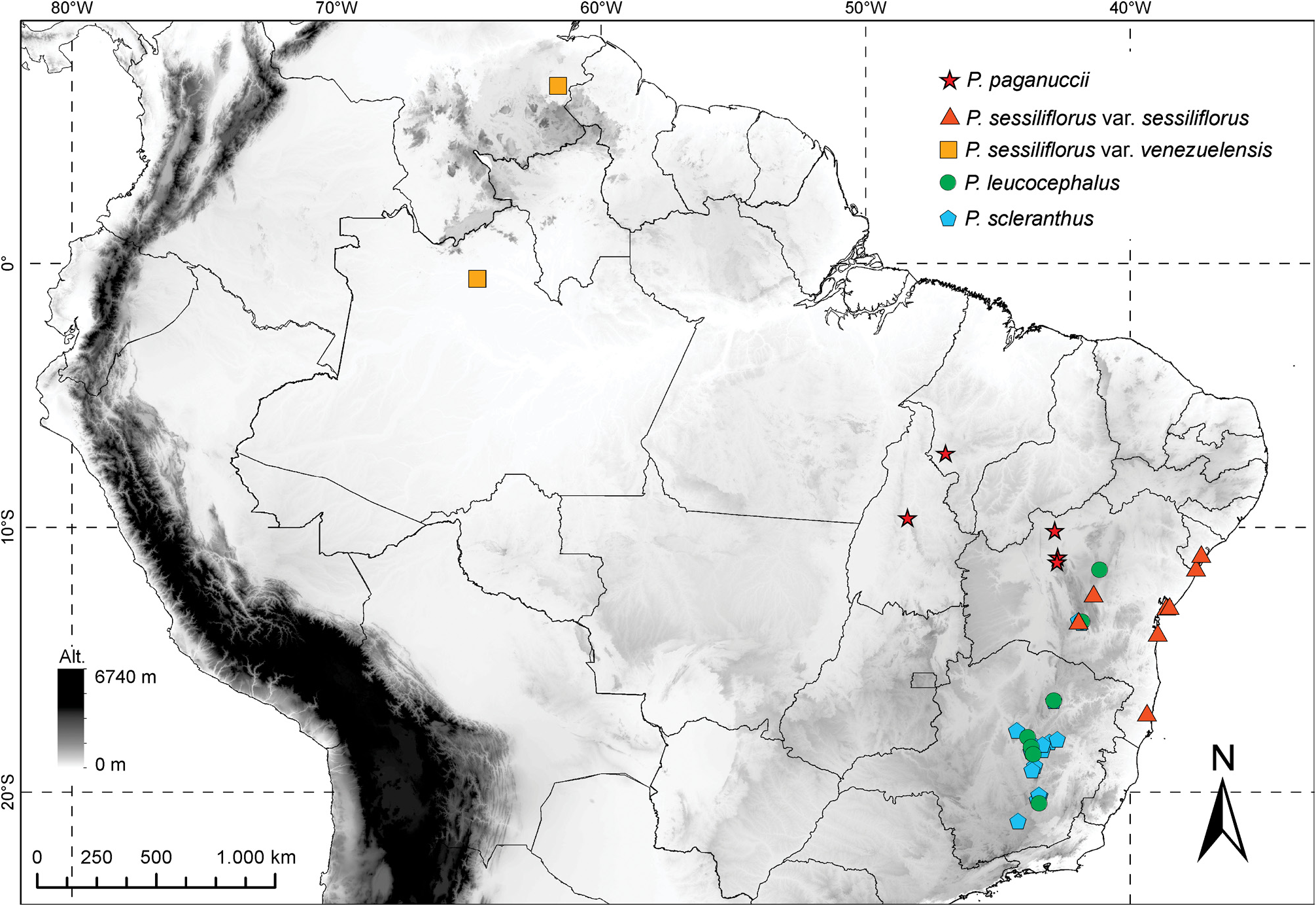
Paepalanthus leucocephalus is sister of P. scleranthus and they are the only species in Paepalanthus subg. Thelxinoë according to Ruhland (1903). They also share the annual caulescent habit of small plants, with the absence of scapes and spathes, capitulum sessile, involucral bracts not differentiated from the apical leaves, floral bracts acuminate, and dimerous flowers. They are sympatric in the Espinhaço Range in some mountains in Minas Gerais, and also in Rio de Contas, Bahia. There is also a population occurring in sandy soils further north in Morro do Chapéu in Bahia ( Figure 3 View FIGURE 3 ). P. leucocephalus is morphologically similar to P. scleranthus ( Figures 6 View FIGURE 6 I-N), sharing also the loss of the nectariferous branches ( Silva et al. 2016), a condition observed in P. paganuccii and P. sessiliflorus as well ( Figures 4–5 View FIGURE4 View FIGURE 5 ). The whitish capitula are a visible character to distinguish P. leucocephalus from P. scleranthus , along with the convex leaves clearly distinguished into sheath and blade, and the morphology of the flowers, illustrated for the first time in this paper. However, the segregation of these species is weak and deserve further investigations as previously stated by Moldenke “The species is extremely close to P. scleranthus and I am not at all certain that the two are distinct” ( Moldenke 1975).
As expected of a widespread species, Paepalanthus leucocephalus encompasses a relevant morphological variation and the relationship with P. scleranthus is complex ( Figures 2 View FIGURE 2 , 4 View FIGURE4 , 6 View FIGURE 6 ). Some populations, as the one represented by the specimen Andrade 625 from Rio de Contas in Bahia, are composed of individuals noteworthy for being smaller than specimens from other populations, especially from those occurring in the Southern Espinhaço Range. The populations from Minas Gerais, occurring mostly in the Diamantina Plateau , also have more hairy floral structures and a shinier white capitulum. The inclusion of specimens from the whole morphological and geographical range of P. leucocephalus and P. scleranthus in future studies at the population level is crucial to circumscribe these species more accurately.
Additional selected material examined:— BRAZIL. Bahia: Morro do Chapéu, Tabuleiro dos Tigres , 11°36’09”S, 41°09’44”W, 20 July 2005, M. J. G GoogleMaps . Andrade et al. 589 ( HUEFS); Rio de Contas, Serra Marsalina , campo cerrado, 16 August 2006, M. J. G . Andrade et al. 620 ( HUEFS); Idem, M. J. G . Andrade et al. 625 ( HUEFS) . Minas Gerais: Datas, 04 km da estrada Datas-Milho Verde , 07 April 2004, M. J. G . Andrade et al. 549 ( HUEFS); Diamantina , 05 May 2010, C . Munhoz et al. 7283 ( UB); Grão Mogol, Trilha do Barão , 02 April 2004, M. J. G . Andrade et al. 481 ( HUEFS); Mariana, Parque Estadual do Itacolomi , 06 April 2018, D. Rodrigues et al. 27 ( OUPR) .


| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| K |
Royal Botanic Gardens |
| M |
Botanische Staatssammlung München |
| J |
University of the Witwatersrand |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| HUEFS |
Universidade Estadual de Feira de Santana |
| C |
University of Copenhagen |
| UB |
Laboratoire de Biostratigraphie |
| OUPR |
Universidade Federal de Ouro Preto, Campus Universitário |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |