
Nemobius interstitialis Barranco, Gilgado & Ortuño, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3691.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:860039CB-3547-4088-B4CE-0DD240559ACA |
|
DOI |
https://doi.org/10.5281/zenodo.5682232 |
|
persistent identifier |
https://treatment.plazi.org/id/03A087AE-FF90-C159-F2CE-FF51FAAF1743 |
|
treatment provided by |
Plazi |
|
scientific name |
Nemobius interstitialis Barranco, Gilgado & Ortuño |
| status |
sp. nov. |
Nemobius interstitialis Barranco, Gilgado & Ortuño sp. nov.
Figures. 2–28 View FIGURES 2 – 3 View FIGURES 4 – 9 View FIGURES 10 – 15 View FIGURES 16 – 22 View FIGURES 23 – 24 View FIGURE 25 View FIGURE 26 View FIGURE 27 View FIGURE 28
Material studied. A total of 2312 specimens (187 males, 281 females and 1744 nymphs) were collected.
Type series. Holotype: 1 male, 11-XI-2012. Paratypes: 5 males and 7 females, 24-XI-2011; 1 male, 27-XII- 2011; 1 male, 11-V-2012; 9 males and 5 females, 2-VIII-2012; 22 males and 18 females, 13-IX-2012; 25 males and 38 females, 11-X- 20122; 1 male and 4 females, 15-XI-2012; All: Puebla de la Sierra, Madrid, Spain. Altitude: 1490 m.a.s.l., mountain scree in the slope of the peak "Cabeza Minga". UTM 30T 4642 4541. Ortuño and Gilgado leg.
The Holotype and 19 Paratypes have been deposited in Museo Nacional de Ciencias Naturales de Madrid, type number 2382. Another 20 Paratypes have been deposited at Dpto. de Ciencias de la Vida in Universidad de Alcalá. The remaining paratypes and specimens are in the collection of the Dpto. Biología y Geología at Universidad de Almería and the Pablo Barranco collection.
Biometry. Biometric comparisons between N. interstitialis sp. nov. and N sylvestris are shown in Table 1 View TABLE 1 . Mean values ??with standard deviations, minimum and maximum values ??for each parameter considered, and the FL/PL and TL/PL index values are shown, as are all statistically significant differences.
N. interstitialis sp. nov. N. sylvestris GLM Diagnosis. A cricket of small size (males 1.05–1.33 cm, females 1.03–1.15 cm). Body pubescent and covered with long chaetiform hairs. Foreleg tibiae lacking a tympanum on either side in both sexes. Male forewing without a stridulatory vein, but with a strongly developed speculum and chordal cells with central ridges around which are profuse secretory pores. Inner margin of female forewing oblique and curved, overlapping in the proximal half ( Figure 3 View FIGURES 2 – 3 ). Male genitalia triangular in shape, with the apical lobe of the epiphallus highly developed.
Male: Head black-brown to black, clypeus and labrum brown. Eyes slightly prominent. Scape dark brown with transverse brownish antennae. Fastigium smooth, a little wider than the scape, with two yellow dots between the median ocellus and the scape, where two pairs of strong setae are inserted. Another three pairs of longitudinal setae in the vertex, from the medium ocellus backward, and another pair just before the inner margin of the lateral ocellus. Space between the median and lateral ocelli a little less than that between the lateral ocelli. Maxillary palps light in color, except for the last segments which are dark at the apices. Third joint cylindrical, one-and-a-half times longer than the fourth, which is conical and flattened toward the apex. Fifth joint more than twice the length of the fourth, conical. Labial palpi light in color with the last segment dark. Basal joint with an oblique upper margin at the level of the second articulation. Submentum with lateral margins rounded ( Figure 4 View FIGURES 4 – 9 ). Mentum trapezoidal. Prementum robust, basal segment of paraglossa not welded to the prementum.
Pronotum transverse, brownish, ornamented with dark dots; fore and hind margins almost straight with long setae. Lateral lobes dark with many strong and long setae, lower margin very slightly sinuous. Forewings blackbrown, short and brachypterous. Right forewing with axillary (Ax), anal node (AN), anal (A), ulnar (U), postaxillary (Pa) and discoidal (D) veins weakly marked. In contrast, the vein prolongations that form the distal cells strongly patent ( Figure 5 View FIGURES 4 – 9 ). Mirror cells [all of medium (M), external (E), and internal (I) cells] and chordal cells (c. c.) very highly developed with a median ridge on each cell ( Figure 5 View FIGURES 4 – 9 ). Concave surfaces surrounding the ridges have many grouped excretory pores; the number of pores per group is variable ( Figure 6 View FIGURES 4 – 9 ). Stridulating vein (SV) of the right forewing absent, stridulatory file with teeth absent ventrally ( Figure 7 View FIGURES 4 – 9 ). SV absent also from left forewing ( Figs. 8 and 9 View FIGURES 4 – 9 ). Hindwings vestigial.
Legs thin, light in color with some dark ornamentation, mostly on the outer side of the hindmost tibia and femur. Femora and tibiae of foreleg and median leg with strong dispersed setae. Foreleg tibia without a tympanum on any side ( Figure 10 View FIGURES 10 – 15 ); with two strong apical spurs, the inner of which is the longer and thicker. Median leg tibia with three apical spurs, one inner and dorsal and two ventral, all of the same size. Tarsomeres 1 of the foreleg and median legs with two serrulated parallel ventral rows of small black spines. Second tarsomere very short with several ventral rows of spines. Thoracic sternites typical of the genus. Hindleg tibiae armored dorsally with three inner and three outer spurs, all strong and thin, arranged alternately ( Figure 11 View FIGURES 10 – 15 ); with three pairs of apical spurs of which those in the middle are shorter and more distal, the proximal pair asymmetric, and the inner pair more developed.
Body dark, covered with dense short setae, predominantly in the pleural zone. Sternites a lighter honey in color. Tenth tergite deeply sloped. Subgenital plate triangular with the apex slightly blunted. Cerci light and hairy with three types of setae of which the long and thin, and short and hard types are located all along the cercus; and globose setae appear on the inner side of the basal part of cercus. In general, all of the surface of the cercus bears many short setae or microtrichia (mt), among which setae termed filiform (f) setae and very thin setae termed tricoid (t) setae are interspersed. Both of the latter types of setae are inserted into pores that are bordered. Globose setae termed club-shaped setae (cs) are located on the inner side of the first half of the cerci; these are smaller basally and increase in size toward the apex ( Figure 12 View FIGURES 10 – 15 ). These cs setae presents an internal border (ir).
Male genitalia ( Figure 14 View FIGURES 10 – 15 ): Short and more triangular in shape than are those of N. sylvestris . Apical lobe of the epiphallus well-developed, broad and dilated, overhanging the margins of the sclerites of the epiphallus. External margin of the dorsal plate of the epiphallus nearly straight and convergent. Basally, the plate slopes at a right angle. Apodemes narrow, almost straight, and increasing progressively in width toward the apex, which is angled at the inner margin. Spermatophore plate well developed.
Female: General shape as in male, but with lighter tergites ornamented with dark dots. Forewings very short, reaching the hind margin of the first abdominal tergite. Six longitudinal parallel veins very patent dorsally ( Figure 13 View FIGURES 10 – 15 ). Lateral lobes with four weakly curved veins. Inner margin of forewings oblique and curved, overlapping in the proximal half. Ovipositor dark and short and does not extend beyond the middle of the extended hindleg tibia, as is characteristic of the genus. Apices of dorsal valves weakly serrulated. Subgenital plate short with a deep obtuse furrow.
Measurements in centimeters (males, females): length of pronotum (0.18–0.25, 0.18–0.24); width of pronotum (0.16–0.23, 0.18–0.26); length of right forewing (0.22–0.36, 0.18–0.24); length of hindleg femur (0.47–0.75, 0.16– 0.23); length of hindleg tibia (0.32–0.50, 0.33–0.51); and ovipositor (0.40–0.54).
Instars. Eight instars have been identified in N. interstitialis sp. nov. They can be readily distinguished on biometrical and morphological bases (with reference to wing pads and ovipositor valves); they match stages described by Gabbutt (1959b) in N. sylvestris . The four first instars are separated only by size, principally hindleg femur length (Gabbutt 1959b; Barranco 2012). Sexes cannot be separated before the fifth instar, when the four ovipositor valve processes can be seen, although wing pads are still not developed. At the sixth instar, wing pads remain undeveloped, but a small ovipositor not extending beyond the end of the body can be seen in females. At the seventh instar, wing pads are present in both sexes, these are laterally arranged and separated dorsally in the middle of the second thoracic segment. In this instar the ovipositor extends beyond the end of the body. At the eighth instar the wing pads are bigger, contact each other dorsally in the middle of the thoracic segment, and the ovipositor is twice the length of that in the prior instar. Table 2 shows the proportions of samples in each instar, separated by sex from the fifth instar, obtained throughout the sampling period.
Etymology. The specific epithet “ interstitialis ” (Latin for interstitial) refers to the lifestyle of the species; it lives among the interstices of stony debris on a mountain scree.
Biology. Dissection of the alimentary canal of 10 adult specimens and 10 last-stage nymphs yielded amorphous fibrous material composed principally of vegetable fibres, leaf fragments, and germinated spores. According to Gabbutt (1959a), N. sylvestris is omnivorous, but feeds principally on leaves.
The location of the pitfall traps, both in the MSS and on the surface, allowed us to obtain information on the population dynamics of N. interstitialis sp. nov. Table 2 shows capture data grouped by date and sex, from both surface and MSS traps, as well as the number of traps in which individuals were caught each month. As shown in Figure 25 View FIGURE 25 , the new species is strongly interstitial in habit; 94.14% of individuals were trapped in the MSS. This is based on a total catch of 2152 individuals, mostly in spring and summer ( Figures 25–27 View FIGURE 25 View FIGURE 26 View FIGURE 27 ). Only 160 individuals were trapped on the surface, suggesting that although N. interstitialis sp. nov. is present all year on the surface, it occurs there only sporadically.
The nymphal instars have been identified and characterized, allowing the age structure of the population to be determined. The population dynamics of each instar are shown in Figure 26 View FIGURE 26 . The new species is univoltine and postembryonic development continues from March to August; instar VI is most common in May, instar VII in June, and instar VIII in July. Adults are the most prevalent stage only in August ( Figure 27 View FIGURE 27 ). Such phenology corresponds exactly with that described for N. sylvestris (Gabbutt 1959a) . According to the classification of cricket cycles by Masaki & Walker (1987) and Masaki (1996), this new species is similar to type “G”. The behavior is intermediate between heterodynamic and homodynamic, because individuals are present all year, lacking any diapausa controlled by photoperiod and temperature, but with a unimodal age structure with the maximum in spring and summer. The population decreases during autumn and winter, although some individuals survive in the MSS by avoiding the frost and snow of the surface. In August and September, a second peak in the numbers of initial postembryonic stages is evident ( Figures 26 View FIGURE 26 and 27 View FIGURE 27 ), probably attributable to an early hatch of eggs placed by the first imagos. However, in October, the population decreases drastically, and the numbers of individuals are greatly reduced in number of during the autumn and winter. Such a dynamic appears to correspond to that described by Gabbutt (1959a) for N. sylvestris , the nymphs of which (if in the fifth instar or over) can survive the winter, though most of the population is in a winter diapausa (thus existing as eggs). Among Iberian crickets, only those adapted to cavernicolous environments, such as Petaloptila (Zapetaloptila) Gorochov & Llorente (Barranco & Amate 2008) or Dolichopoda Bolívar (Carchini et al. 1989; 1991; 1995; Rampini et al. 1983) have homodynamic cycles. Such cycles are multimodal; permanent populations with all stages can be found all year, because of the stable and constant environments of the caverns.
An interesting feature of the population is the predominance of females in most samples ( Figure 28 View FIGURE 28 ): 1077 females and 943 males were counted (including crickets at instar V or above, and adults). This equates to 53.32% females and 46.48% males. This relationship differs slightly from that calculated by Gabbutt (1959a) for N. sylvestris ; the populations of which have equal numbers of males and females.
Two of the most important abiotic factors influencing species activity are temperature and humidity, which we measured in our study. As is true of other MSSs (Pipan et al. 2011), our data show daily and seasonal variations in temperature, but these are much less than at the surface ( Figure 29 View FIGURE 29 A, B). For example, the MSS experiences daily variations of about 2 degrees, whereas, outside, the variation is 15 or 20 degrees. In terms of humidity, this is higher in the MSS than on the surface and only in summer does it decrease to 30% from time-to-time, such low humidity is often noted outside ( Figure 29 View FIGURE 29 A, B). Very probably, these features favor species activity in the interstices of the scree during summer, where the humidity is higher and the temperature oscillates between 15ºC and 20ºC, in contrast to the range (10ºC to 30ºC) on the surface. However, it is also surprising that sporadic catches of some individuals were made in the coldest months when the minimum MSS temperature reached –3ºC (on the surface it attained –10ºC) and the maximum barely reached 5ºC.
In terms of accompanying fauna, N. interstitialis sp. nov. shares its habitat with a number of species that are predominantly epigean. The presence of such fauna in the MSS may be explained by the porosity of the scree, right up to the surface, but also by the temperature and humidity conditions in the scree ( Figure 29 View FIGURE 29 ). Predation of N. interstitialis sp. nov. may not impose strong pressure on the species; the population is numerous. Also, few species of predators were noted among the accompanying fauna, and these were not very numerous. Some small species might prey on the nymphs of N. interstitialis sp. nov., these might include some Carabidae such as Trechus schaufussi Putzeys , Platyderus montanellus Graells , Pseudomasoreus canigoulensis (Fairmaire & Laboulbène) , and Nebria salina Fairmaire & Laboulbène ; the Formicidae Camponotus pilicornis Roger , and Camponotus cruentatus Latreille ; and several species of Staphylinidae, Opiliones, Pseudoscorpiones , and Araneae which move inside the interstices of the scree. Some predators on late-stage nymphs and imagos may be the Dysderidae Harpactocrates gurdus Simon , a spider well represented in the MSS; some Chilopoda of the genera Lithobius Leach ; and Scolopendra cingulata Latreille. Some vertebrates have also been observed in the scree; these may also be predators of the new species. The vertebrates include the Lacertidae Podarcis hispanica (Steindachner) and the Insectivora Sorex minutus Linnaeus. Other non-predators sharing the habitat with Nemobius interstitialis sp. nov. are Coleoptera of the families Ptinidae , Cryptophagidae , Leiodidae and a few Lagriidae . It is also noteworthy that larvae of Lampyridae , mostly of the genus Lampyris Geoffroy , and a small number of Nyctophyla reichii Jacquelin du Val, were present during almost the entire year of sampling. Other orders of insects, such as Diptera , were also present in high numbers, most of which belonged to the genera Megaselia Rondani and Drapetis Meigen.


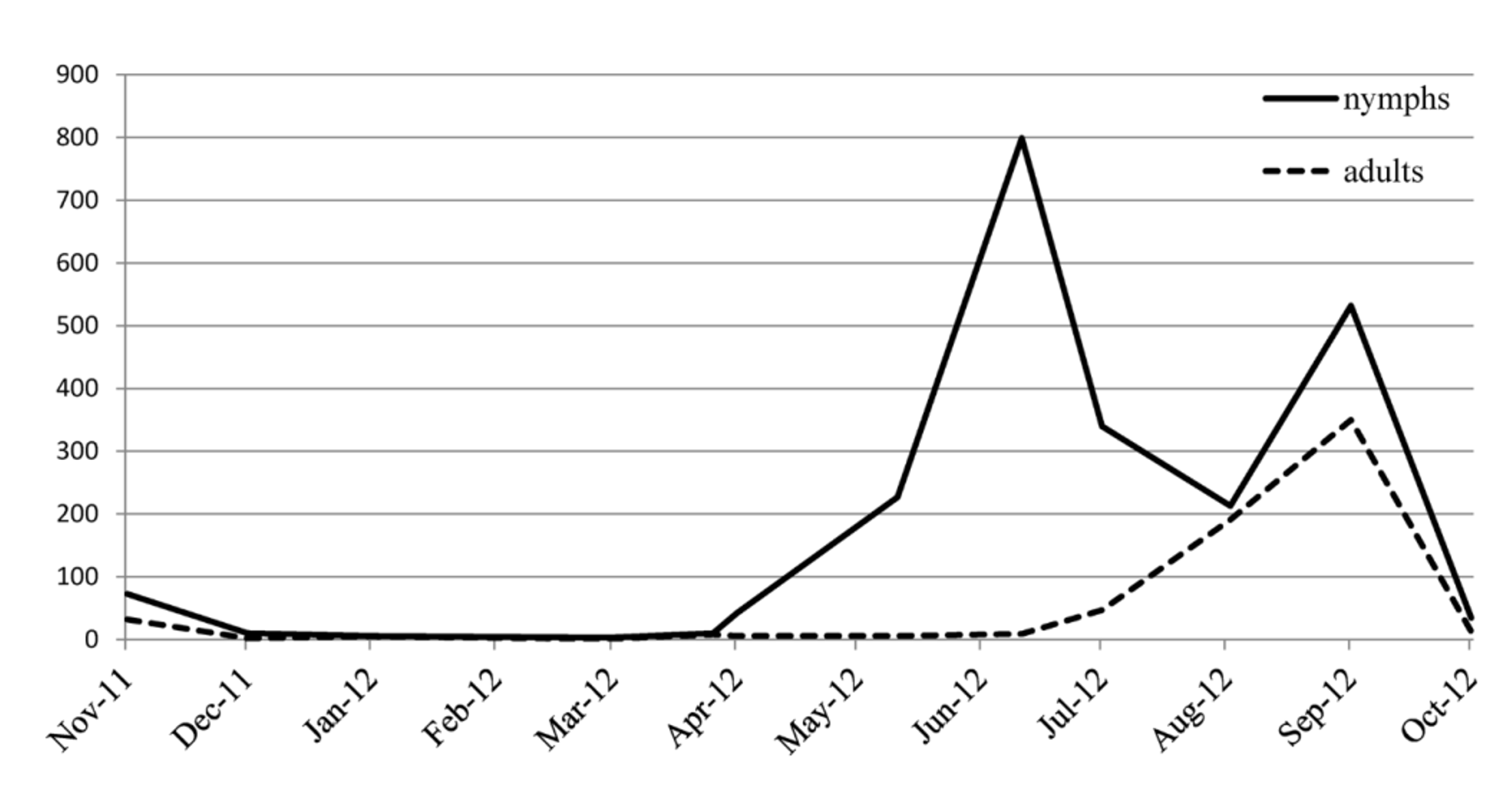




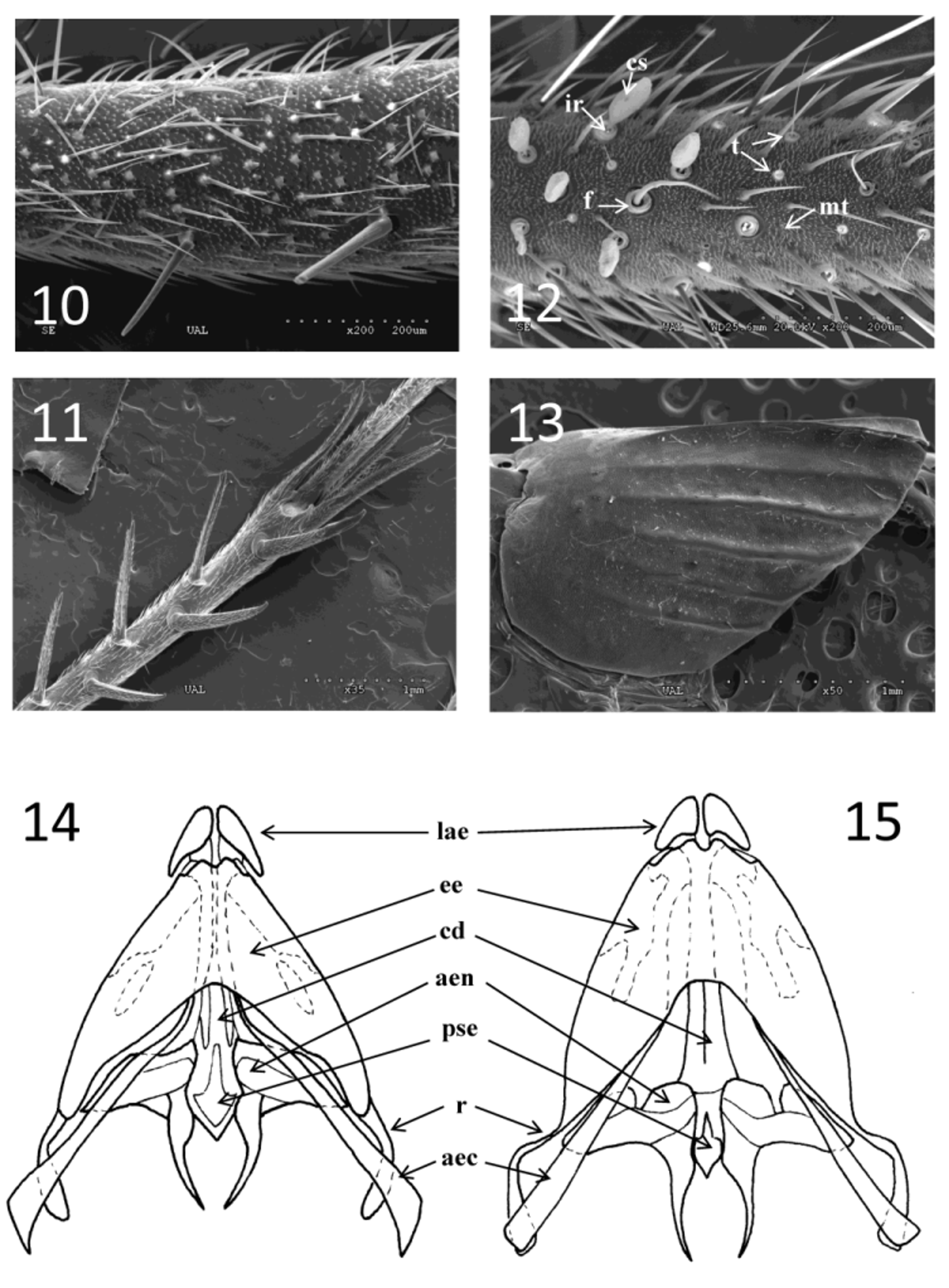


TABLE 1. Measured parameters in mm. Intervals of variation and average values (with standard deviations) are shown. Right column: Statistical analysis (General Linear Model), p: significance, F: Fisher’s statistical parameter.
| Males n= 23 | Females n= 42 | Males n= 23 | Females n= 22 | Males p, F | Females p, F |
|---|---|---|---|---|---|
| Pronotum length (PL) 0.16-0.23 0.21 ± 0.02 | 0.18-0.24 0.21 ± 0.01 | 0.13-0.19 0.17 ± 0.01 | 0.17-0.21 0.20 ± 0.01 | 0.0000 37.51 | 0.0048 8.57 |
| Pronotum width (PW) 0.16-0.23 0.21 ± 0.02 | 0.18-0.29 0.26 ± 0.02 | 0.22-0.28 0.26 ± 0.02 | 0.23-0.30 0.27 ± 0.03 | 0.0370 4.63 | 0.0005 13.65 |
| Fore wing length (TL) 0.22-0.36 0.30 ± 0.04 | 0.15-0.23 0.20 ± 0.02 | 0.32-0.42 0.37 ± 0.02 | 0.18-0.23 0.20 ± 0.02 | 0.0000 66.91 | 0.0000 26.25 |
| Hind femur length (FL) 0.47-0.75 0.63 ± 0.08 | 0.56-0.75 0.69 ± 0.04 | 0.49-0.67 0.60 ± 0.05 | 0.55-0.78 0.63 ± 0.07 | 0.0047 8.87 | 0.0523 3.91 |
| Hind tibia length (TL) 0.32-0.50 0.43 ± 0.05 | 0.33-0.51 0.43 ± 0.04 | 0.36-0.45 0.40 ± 0.03 | 0.38-0.49 0.44 ± 0.05 | 0.0012 11.92 | 0.0598 3.67 |
| Ovipositor | 0.40-0.56 0.47 ± 0.04 | 0.54-0.68 0.61 ± 0.04 | 0.0000 90.49 | ||
| FL/PL 2.68-4.00 3.25 ± 0.23 | 2.79-3.83 3.27 ± 0.26 | 3.06-4.69 3.55 ± 0.18 | 2.71-3.90 3.20 ± 0.33 | 0.0087 7.57 | 0.5501 0.36 |
| TL/PL 1.76-2.56 2.19 ± 0.18 | 0.71-1.06 0.95 ± 0.10 | 2.05-3.00 2.34 ± 0.12 | 1.90-2.45 2.21 ± 0.16 | 0.0162 6.25 | 0.0000 463.61 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.